
Береза повислая . Широко распространена в европейской части нашей страны от южных районов до границ лесотундры.
Обычно она участвует в качестве примеси к широколиственным или хвойным породам в самом разном сочетании в различных типах лесорастительных условий. Несмотря на ее широкое распространение, морфология корневой системы березы до последнего времени малоизучена.
В условиях свежей су дубравы на средне-дернованных среднеподзолистых почвах в сосново-березовых насаждениях I класса бонитета, полнотой 0,8 корни березы имеют 10-15 хорошо развитых горизонтальных корней первого порядка, образующих в верхних горизонтах почвы густую сеть скелетных, полускелетных и всасывающих корней. В 27-летнем возрасте длина горизонтальных корней первого порядка достигает 8,05 м, диаметр 13,6 см. Стержневые корни развиты слабо, проникают в глубь почвы на 95-115 см. У большинства деревьев наблюдается большое количество хорошо развитых вертикальных ответвлений от горизонтальных корней, некоторые из них проникают в почву на большую глубину по сравнению со стержневыми корнями. Однако у отдельных деревьев отмечается полное отсутствие вертикальных ответвлений. Протяженность скелетных корней горизонтальной ориентации и их ответвлений зависит от группы роста дерева.
По сравнению с другими породами береза имеет самый высокий коэффициент ветвистости — 17,2 (сосна 3,0, дуб 1,5, ель 5,6, клен 1,8). Площадь проекции корневых систем составляет 33,1-46 м 2 , объем почвы, занимаемый корнями, 11,0-43,7 м 3 в зависимости от группы роста дерева.
Интенсивность корненаселенности занимаемого объема почвы у разных деревьев различна - от 19,1 до 111,1 м/м 3 , т. е. по сравнению с сосной в 1,8-2,6 раза больше. Среднегодовой прирост по объему почвы, занимаемому корнями, достигает 4,1 м 3 , по общей длине корней 15,4 м 3 , по поверхности всасывающего пространства корней 9,1 дм 3 .
Граб обыкновенный . Обычно в естественных условиях граб не образует чистых древостоев, однако значение его как сопутствующей породы велико. Широко распространен в широколиственных лесах юго-запада европейской части СССР. Характеризуется как порода, имеющая мощную поверхностную корневую систему.
В корневой системе граба в 15-летних дубово-грабовых культурах в условиях Винницкой области на серых лесных почвах (тип лесорастительных условий - свежая дубрава) преобладают корни горизонтальной ориентации.
Однако в этом же возрасте часто обнаруживаются хорошо развитые стержневые корни, имеющие высокую степень разветвленности и проникающие на глубину до 1,9 м. Горизонтальные корни первого порядка достигают длины 5,9 м. Степень разветвленности высокая, имеются скелетные корни седьмого-восьмого порядков ветвления. В общей протяженности корней преобладают корни второго порядка ветвления, в общей массе - первого, а по количеству ответвлений - третьего порядка ветвления.
Бук лесной . На территории СССР бук естественно произрастает в Калининградской области, в Карпатах и Предкарпатье, Кодрах Молдавии и в Крыму. Строение корневой системы бука лесного, так же как и пихты белой, изучено недостаточно.
Так же как и у ели, корневая система бука в 11-22-летних елово-буково-пихтовых культурах в условиях Карпат на высоте 750-1000 м над ур. м. имеет выраженное поверхностное строение.. Стержневой корень в 11-22-летнем возрасте деревьев обычно отсутствует. Он трансформируется в короткое утолщение, являющееся продолжением ствола дерева.
Относительное участие горизонтальных корней в общей длине скелетных составляет 99,2-99,96 %, в общей массе корневой системы 70,1-73,2 %. У отдельных деревьев может быть по 3-4 вертикальных ответвления, некоторые из которых имеют интенсивное ветвление и проникают в глубь почвы по расщелинам до 160 см. Однако на более глубоких щебенистых почвах встречаются единичные деревья, стержневой корень которых в 18-летнем возрасте через расщелины в скальных породах проникает до глубины 241 см.
Корни бука первого порядка отличаются сильной сбежистостью у основания. Затем на расстоянии 0,1 длины их диаметры уменьшаются относительно длины более умеренно, и корни приобретают более выраженную шнуровидную форму. Характер сбежистости корня выражают следующие коэффициенты формы по относительным длинам: 0,1-62,3; 0,2-50,4; 0,5-27,8; 0,7-16,5; 0,9-7,9 %. Коэффициенты формы и коэффициент объема корня (0,1800) свидетельствуют об относительно небольшой сбежистости скелетных корней бука.
Площадь проекции корневой системы у деревьев лучшего роста в 22-летнем возрасте составляет 60,6 м 2 (у средних деревьев 21,2, у отстающих в росте 10,5 м 2). Объем почвенного пространства, занимаемого корневыми системами в этом возрасте, у деревьев лучшего роста равен 36,4 м 3 , средних 12,7, отстающих в росте 3,2 м 3 . Коэффициент компактности корневой системы соответственно составляет 14,3; 16,6 и 20,6 м/м. Эти показатели несколько выше, чем у ели европейской.
Дуб черешчатый . Произрастает в пределах своего естественного ареала в средней и южной зонах европейской части СССР, в Крыму и на Кавказе. В пределах такого широкого ареала дуб встречается в различных типах лесорастительных условий и типах леса. Являясь породой, требовательной к плодородию почвы, дуб в естественных условиях образует смешанные насаждения на почвах, представляющих относительно широкий диапазон как по плодородию, так и по характеру увлажнения. Однако в определенных эдафических условиях он может иметь III-IV классы бонитета, образуя второй ярус в смешанных древостоях на сухих, бедных песчаных почвах. В более благоприятных условиях он выходит в первый ярус, достигая II или I класса бонитета, а на богатых, хорошо увлажненных почвах - I и Iа классов бонитета.
Лучшие условия для роста дуба черешчатого - свежие и влажные серые лесные суглинки, деградированные черноземы, мощные бурые горно-лесные почвы. Под влиянием почвенных условий формируются особенности строения корневой системы дуба. Обладая способностью образовывать с самых первых лет мощный стержневой корень, дуб на почвах с избыточным увлажнением формирует поверхностную корневую систему, при относительно недостаточном увлажнении дает хорошо развитые вертикальные ответвления от горизонтальных корней, на почвах с наличием уплотненных горизонтов типа ортштейна образует второй ярус корней над его поверхностью.
В корневой системе дуба на черноземах обыкновенных малогумусных с признаками южного чернозема в дубово-ясеневых культурах преобладают корни вертикальной ориентации. Вертикальные ответвления начинают появляться в 10-летнем возрасте, но уже в 18 лет они составляют около 20 % общей длины горизонтальных корней. Горизонтальные корни слабо разветвлены. Наиболее высокое относительное участие составляют скелетные корни первого порядка. Разветвление стержневого корня более интенсивное по сравнению с горизонтальными корнями.
Глубина проникновения стержневых корней дуба достигает в 10-летнем возрасте 4,05, в 18-летнем 4,86. Развитие вертикальных ответвлений от горизонтальный корней интенсивное. Некоторые из них по диаметру и длине превышают стержневой корень, достигая глубины 250-280 см. Основное количество корней располагается в верхних горизонтах почвы. У деревьев лучшего роста до 83,8 % горизонтальных скелетных корней размещается на глубине до 20 см, 95 % - в 0-40-сантиметровом слое почвы.
На черноземах деградированных формируется более поверхностная корневая система дуба. Относительное участие корней горизонтальной ориентации на 13-20 % больше с соответственным уменьшением количества вертикальных ответвлений и стержневых корней. В то же время отмечается значительно большая разветвленность горизонтальных и стержневых корней, несмотря на то что наибольшее относительное участие сохраняется за корнями первого порядка ветвления. Резко сокращается глубина проникновения стержневого корня в почву. Она составляет у деревьев лучшего роста в 9-летнем возрасте 167 см, 16-летнем 183, 18-летнем 195 см. Это более чем в 2 раза меньше по сравнению с глубиной проникновения корней дуба черешчатого на черноземах обыкновенных малогумусных в южной степи.
На серых лесных почвах в свежих типах лесорастительных условий в структуре корневой системы дуба относительное участие вертикальных ответвлений в 2-2,5 раза больше, чем на деградированных почвах, и почти в 3 раза больше, чем на черноземах обыкновенных. Интенсивность развития горизонтальных корней и стержневого корня здесь значительно выше, чем на деградированных и обыкновенных черноземах. Общая протяженность скелетных корней в этих условиях уже в 10-летнем возрасте в несколько раз больше, чем у деревьев 16-19-летнего возраста в ранее рассмотренных условиях. Глубина проникновения стержневого корня на серых лесных почвах достигает в 10-летнем возрасте 190 см, а в 25-летнем 555 см, что намного больше по сравнению с другими почвенными разностями. Вертикальные ответвления также интенсивно развиты и уже в 10-летнем возрасте достигают глубины 215 см. Следовательно, свежие лесные суглинистые почвы наиболее благоприятны для роста дуба черешчатого.
Некоторые изменения в строении корневых систем по сравнению с серыми лесными почвами Правобережья Украины и черноземами наблюдаются в условиях дерново-подзолистых и серых лесных почв западной части лесостепи. Общая интенсивность развития корневых систем здесь меньше, чем на черноземах и серых лесных почвах Правобережья. Намного слабее развивается стержневой корень, росту которого в глубь почвы здесь препятствуют уплотненные горизонты почвы ортштейнового характера с признаками оглеения. Глубина проникновения стержневого корня достигает на дерново-подзолистых почвах в 14-летнем возрасте 160 см, на серых лесных 220 см.
Корневые системы деревьев старшего возраста полностью оформляются на последнем этапе своего развития. Горизонтальные корни дуба в 90-летнем возрасте (насаждение I класса бонитета, почва средне-дерново-слабоподзолистая легкосупесчаная на флювиогляциальных отложениях) представлены мощными корнями первого порядка, расположенными у поверхности почвы на глубине до 30 см. От стержневого корня на глубине 32-60 см отходят 11 корней горизонтальной ориентации.
Интенсивность ветвления корней довольно слабая, наибольшее количество ответвлений-тратьего порядка. Горизонтальные корни образуют сеть шнуровидных корней, расположенных у самой поверхности почвы. Длина наиболее развитого корня первого порядка составляет 22,4 м. Общая длина скелетных поверхностных горизонтальных корней с ответвлениями до пятого порядка составляет 1995 м, глубинных 207,9 м. Поверхностные горизонтальные корни имеют вертикальные ответвления длиной до 113 м, что составляют около 5 % общей протяженности этих корней. Глубинные корни горизонтальной ориентации отличаются слабым развитием. Наибольшее относительное участие у этой категории корней составляют корни второго порядка.
Особенностью глубинных корней горизонтальной ориентации является их способность образовывать вертикальные ответвления, которые могут быть направлены не только вглубь, но и вверх - к поверхности почвы. Поверхностные горизонтальные корни обладают хорошо выраженной досковидностью, у основания корня их вертикальный диаметр может превышать горизонтальный в 5-8,5 раза. Различие в размерах вертикального и горизонтального диаметров исчезает на расстоянии 60-140 см от основания корня в зависимости от его крупности. Глубинные горизонтальные корни досковидностью не обладают.
Стержневой корень 90-летнего дуба имеет множество ветвлений большого диаметра, сильно переплетенных между собой и почти полностью сросшихся в верхней части. Со стержневым корнем и его ответвлениями переплетаются и срастаются якорные корни, образовавшиеся в непосредственной близости у ствола дерева. Глубина проникновения основного корня составляет 178 см, якорных - до 250 см. Стержневая часть корневой системы дуба черешчатого представляет единое, монолитное сплетение стержневого корня и ближайших якорных корней, сросшихся между собой.
Протяженность поддающихся учету элементов этой сросшейся системы равна 17,8 м. Суммарно протяженность стержневой части корневой системы и вертикальных ответвлений от стержневых корней составляет примерно 130 м, или 5 % общей длины скелетных корней.
У дуба, как и у других древесных пород, в корневой системе в основном участвуют корни горизонтальной ориентации, наиболее интенсивно осваивающие верхние горизонты почвы в пределах 0-60 см. Вместе с тем дуб обладает способностью в оптимальных для него почвенно-гидрологических условиях образовывать глубоко идущий и сильно развитый стержневой корень. Способность же его образовывать вертикальные ответвления от горизонтальных корней несколько меньшая, чем у многих древесных пород (сосна, орех, каштан, липа, ель). Интенсивность разветвления корней дуба слабая, причем значительного варьирования этого признака под влиянием почвенных условий не установлено.
Средний коэффициент ветвистости корней дуба выражается показателем 1,46, что ниже его значения, полученного для других древесных пород. Интенсивность сбежистости скелетных корней дуба определяется коэффициентами формы корня на относительных длинах: 0,1 - 72,4±0,55; 0,2 — 56,2±0,63; 0,5 — 29,8±0,54; 0,7 — 16,7±0,4; 0,9-7,4+0,20. Коэффициент объема горизонтальных скелетных корней дуба 0,1851, что свидетельствует о большой шнуровидности его корней по сравнению с другими породами.
Площади проекции корневых систем дуба могут достигать к 19-летнему возрасту 50 м 2 , к 25-летнему более 60 м 2 . Превышение площади проекций корневых систем над площадью проекций крон составляет от 5,4 до 8,4. Большая глубина проникновения корней в почву обеспечивает дубу быстрое освоение значительных объемов почвенного пространства, что делает исключительным показатель компактности его корневой системы, находящийся в пределах 1,9-10,8.
Липа крупнолистная . Широко распространена в лесах европейской части СССР. Произрастает на различных почвах, предпочитая более богатые, свежие лесорастительные условия. Выступает в естественных насаждениях и используется в культурах в качестве сопутствующей породы с дубом, сосной, лиственницей, как правило, образует второй ярус, а в менее благоприятных условиях - третий.
Корневая система хорошо развита. В ее структуре (в 12-летних культурах, на серых лесных суглинистых почвах) корни горизонтальной ориентации составляют 78,6-93,6 %. У дерева лучшего роста стержневой корень отсутствует, у среднего и отстающего в росте он занимает 3,1 и 9,9 % общей длины скелетных корней. Разветвленность скелетных корней ограничивается образованием корней третьего порядка. Вертикальные ответвления от горизонтальных корней составляют 3,6-11,2 %.
Структура и строение корневой системы липы свидетельствуют о ее поверхностном размещении. Глубина проникновения корней составляет у дерева лучшего роста 40 см за счет углубления горизонтальных корней. В этом 40-сантиметровом слое почвы размещены все 100 % корней дерева лучшего роста. Стержневые корни деревьев среднего роста и отстающих в росте достигают глубины 80 и 70 см. Среднегодовой прирост наиболее крупного горизонтального корня составляет 21,7, среднего 14,3 см. Эти показатели значительно ниже, чем у других древесных пород в этом насаждении (у клена явора 40,8 и 15,7, березы повислой 35,4 и 27,1, сосны обыкновенной 0,43 и 16,3, дуба черешчатого 28,9 и 17,5 см).
Интенсивность разветвленности корней липы средняя. Она характеризуется коэффициентом ветвистости 2,1. Это несколько больше, чем у клена явора (1,8) и дуба (1,5), но значительно меньше, чем у других совместно произрастающих пород (у сосны 2,5, березы 17,2).
Сбежистость корней липы определяется коэффициентами формы на относительных длинах: 0,1 - 0,657±0,016; 0,2 - 0,472±0,017; 0,5 - 0,330±0,018; 0,7 - 0,220±0,012; 0,9 - 0,104±0,04. Коэффициент объема корней 0,1701, что соответствует среднему значению среди других древесных пород.
Площадь проекции корневой системы липы меньше, чем у других пород: у дерева лучшего роста 9,3 м 2 , среднего 10,0, отстающего в росте 1,3 м 2 . Объем почвенного пространства, занимаемого корневой системой, соответственно равен 2,2; 2,7; 0,3 м 3 . Коэффициент компактности корневой системы очень высок. У дерева лучшего роста он составляет 37,7, у среднего 19,1.
Клен остролистный . Как и дуб, клен широко распространен в лесах европейской части СССР. Однако биометрическая характеристика корневой системы клена малоизучена. При совместном произрастании в культурах дуба клен остролистный имеет хорошо развитую корневую систему, состоящую из стержневого корня, проникающего в глубь почвы на 3 м, и мощных корней горизонтальной ориентации. Интенсивность корненаселенности верхних горизонтов почвы у клена почти не уступает таковой у дуба черешчатого.
Клен явор . Произрастает в Карпатах в качестве примеси в еловых, буковых и пихтовых лесах. Вместе с тем он широко распространен в равнинных лесах Прикарпатья. Древесина этой породы имеет большую хозяйственную ценность.
Клен явор отличается хорошо развитыми стержневым и горизонтальным корнями. Горизонтальные корни имеют ответвления третьего и четвертого порядков. Корни вертикальной ориентации представлены стержневым корнем и его ответвлениями второго и третьего порядков. Основная масса корней на бурых горно-лесных почвах расположена в 0-30-сантиметровом слое, однако отдельные стержневые корни проникают на глубину более 1 м. У явора, как и у других пород, в общей длине корней преобладают корни горизонтальной ориентации (81,2-99,2 %), участие же стержневого корня в общей массе корней у явора значительно больше. В общей протяженности корней явора преобладают корни второго-третьего порядков.
Протяженность скелетных корней у явора меньше, чем у ели, пихты и бука, но в отличие от них у этой породы интенсивнее развиты стержневые корни и вертикальные ответвления от горизонтальных корней.
Различие в почвенных условиях отражается на структуре и строении корневых систем. На глубоких серых лесных почвах относительное участие стержневого корня в общей длине существенно больше, чем на бурых горно-лесных почвах средней мощности, а длина стержневого корня с ответвлениями на серых лесных почвах в 2,5-8 раз больше. В этих условиях более интенсивно развиты и вертикальные ответвления от горизонтальных корней. Максимальная глубина проникновения стержневого корня на бурых горно-лесных почвах составляет 120 см, на серых лесных в 12-летнем возрасте у деревьев среднего роста - 123 см, лучших - 510 см.
Сбежистость горизонтальных корней клена явора характеризуется следующими коэффициентами формы по относительным длинам корня: 0,1 — 67,3±0,01; 0,2 — 46,0+0,01; 0,5 — 24,4±0,07; 0,7 — 16,2+0,01; 0,9 - 9,2±0,003. Коэффициент объема для горизонтальных корней первого порядка 0,1444. По интенсивности сбежистости корни явора занимают среднее положение среди представленных в данном учебнике древесных пород. Интенсивность разветвленности корней клена явора очень низкая (средний коэффициент ветвистости составляет 1,8).
Максимальная величина среднегодового прироста по длине горизонтального корня первого порядка составляет 21,7 см, средний прирост 14,8 см, среднегодовой прирост стержневого корня 6,7 см. Отношение интенсивности роста стержневого и среднего горизонтального корней составляет на серых лесных почвах 0,47.
Площадь проекций корневой системы к 18-летнему возрасту на бурых горно-лесных почвах достигает 20,4 м 2 , на серых лесных почвах в 12-летнем возрасте 11,2 м 2 , что соответствует площади проекции корней деревьев 14-летнего возраста (11,5 м 2) на бурых почвах Карпат.
Корневая система клена явора на серых лесных глубоких почвах отличается невысокой компактностью. Благодаря стержневому корню, который интенсивно проникает вглубь, корневая система относительно быстро занимает обширный объем почвенного пространства. В 12-летнем возрасте в этих условиях объем почвы, занимаемый корневыми системами, составляет для дерева лучшего роста 19,3 м 3 , среднего 18,9 и отстающего 1,1 м 3 ; коэффициент компактности корневых систем соответственно 2,6; 2,9 и 2,9 м/м 3 . Однако этот показатель возрастает в 10 и более раз на среднемощных бурых горно-лесных почвах, где у деревьев лучшего роста в 8-летнем возрасте он составляет 36,3, в 12-летнем 26,3 и в 17-летнем 23,2 м/м 3 .
Орех грецкий . В естественных лесах произрастает в горных районах Киргизии. Широко культивируется в Средней Азии, на Кавказе, Украине, в Молдавии и на юге Белоруссии. Предпочитает свежие и влажные достаточно богатые почвы (черноземы и серые лесные почвы). Уже к 6-летнему возрасту на серых лесных почвах у ореха формируются не только хорошо развитые стержневой и горизонтальные корни, но и значительное количество вертикальных ответвлений. Глубина проникновения стержневых корней в этом возрасте составляет в зависимости от группы роста дерева 273, 241 и 194 см. Ответвления от стержневого корня расположены равномерно по всей его длине. Суммарная протяженность вертикальных ответвлений от корней горизонтальной ориентации составляет 6,9-12,3 % общей длины скелетных корней. У каждого дерева насчитывается 8-10 вертикальных ответвлений. Глубина проникновения их также различна в зависимости от группы роста дерева. Так, у деревьев, отстающих в росте, она составляет 49-67 см, деревьев среднего роста 82-124, лучшего 120-241 см. Среднегодовой прирост по длине корней горизонтальной ориентации составляет 61-73 см, по диаметру 3,4-9,5 мм.
Разветвленность корней довольно интенсивная: 420-820 скелетных ответвлений. Наивысший порядок ветвления в этом возрасте - четвертый, однако корней этого порядка очень мало (0,3-0,9 %). Основное относительное участие в общей протяженности скелетных корней принимают корни второго порядка ветвления (39,1-55,8 %).
В общей протяженности скелетных корней ореха грецкого существенное относительное участие принимают вертикальные ответвления от горизонтальных корней. Компактность корневых систем незначительная.
Орех грецкий отличается высокий интенсивностью прироста корней по диаметру, достигающего у корней горизонтальной ориентации 0,95, а у стержневых корней 1,05 см. Площадь горизонтальной проекции корней составляет соответственно по группам роста деревьев 38, 26 и 23 м 2 , что превышает площадь проекций крон соответственно в 2,9; 3,9 и 5,5 раза.
Сбежистость корней характеризуется следующими коэффициентами формы корней на соответствующих относительных длинах: 0,1 - 56,5; 0,2 - 35,1; 0,5 - 26,1; 0,7 - 18,7; 0,9 - 11,4. Коэффициент объема корней 0,1207.
Лещина обыкновенная . Широко распространена в европейской части СССР как подлесочная порода. В пределах своего естественного ареала она встречается в свежих и влажных гигротопах на черноземных, буроземных, серых лесных, дерново-подзолистых почвах, обладающих высоким плодородием.
Корневая система лещины в условиях запада Украины на дерновослабоподзолистых легкосуглинистых почвах в свежей грабовой судубраве в 90-летних сосняках Iа класса бонитета такова: стержневые корни отсутствуют, горизонтальные имеют высокую ветвистость. Общая протяженность скелетных корней одного куста достигает 256 м, из которых корни первого порядка ветвления составляют 8,7, второго 40,8 и третьего 50,5 %. Общее число ответвлений у наиболее развитых кустов 850, в том числе первого порядка 1,1, второго 21,9, третьего 77,1 %. Коэффициент ветвистости корней высокий - 7,8. Сбежистость корней первого порядка характеризуется коэффициентами формы на относительных длинах: 0,1- 0,54; 0,2 - 0,38; 0,5 — 0,25; 0,7 — 0,174 и 0,9 - 0,14. Коэффициент объема скелетных корней первого порядка 0,1224.
Основная масса корней лещины расположена на глубине 0-30 см, однако отдельные корни проникают вглубь до 60 см. Корни лещины, расходясь далеко в стороны от куста, осваивают значительную площадь питания, достигающую 15 м 2 . Несмотря на это, коэффициент компактности корневой системы у нее составляет 28,3 %. Таким образом, корневая система лещины довольно интенсивно населяет верхний горизонт почвы в насаждениях.
Каштан съедобный (посевной) . Каштан съедобный (посевной), европейский, или благородный, естественно произрастает на Кавказе, а также широко распространен в Карпатах, образуя в культурах высокопроизводительные ценные древостой. Каштан образует глубокую корневую систему за счет крупных корней, идущих косовертикально в глубь почвы. Стержневой корень отсутствует. Корневые системы деревьев 10-летнего возраста в культурах каштана с участием дуба черешчатого на карпатских бурых горно-лесных почвах состоят из стержневого корня, горизонтальных корней и вертикальных ответвлений от горизонтальных корней. Часть горизонтальных корней уходят в почву под хорошо выраженным углом в косовертикальном направлении. Скелетных корней первого порядка у деревьев лучшего роста немного, а у деревьев, отстающих в росте, их значительно больше. В то же время у деревьев, отстающих в росте, отсутствуют ответвления второго порядка и вертикальные ответвления от горизонтальных корней, а стержневые корни развиты значительно слабее. Это свидетельствует о том, что более слабые деревья осваивают жизненное почвенное пространство более молодыми горизонтальными корнями первого порядка.
В структуре корневой системы каштана посевного основное относительное участие принимают корни горизонтальной ориентации. Однако обращает на себя внимание очень высокое относительное участие корней вертикальной ориентации у деревьев лучшего и среднего роста. Так, суммарная длина стержневых корней и вертикальных ответвлений составляет у деревьев лучшего роста 25,7, среднего 12,7 %.
В строении корневой системы каштана посевного в 10-летнем возрасте преобладают корни второго порядка ветвления. Так, у дерева лучшего роста горизонтальные корни первого порядка составляют 21,7 %, второго 46,7, третьего 10,9, вертикальные ответвления 15,8 %, стержневой корень с ответвлениями первого и второго порядков 4,9 % общей длины скелетных корней.
Стержневые корни каштана проникают на глубину до 3 м. Вместе с тем глубина проникновения стержневых корней дуба черешчатого при совместном произрастании с каштаном составляет 4,2 м.
Так же, как и у других древесных пород, площадь проекций корневых систем каштана значительно превосходит площадь проекции крон. Это положение характеризуется следующими показателями: у дерева лучшего роста площадь проекции кроны составляет 3,14 м 2 , площадь проекции корней 22,04 м 2 , т. е. в 7 раз больше; у дерева среднего роста соответственно 1,76 и 12,6 м 2 , т. е. в 7,2 раза больше.
Интенсивность населенности почвенного пространства скелетными корнями у дерева лучшего роста 6,7, среднего 6,1, отстающего в росте 13,9 м/м 3 .
Горизонтальные корни каштана посевного относительно малосбежистые. Диаметр корней первого порядка на 0,5 относительной длины составляет 34,9 %, что значительно выше, чем у многих древесных пород. Так же, как и у других пород, горизонтальные корни второго порядка у каштана посевного менее сбежисты, чем корни первого порядка.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter .
Страница 2 из 4
Строение, рост, развитие органов виноградного растения и их функции
К вегетативным органам виноградного растения относятся корень, стебель и листья, к генеративным (репродуктивным) - цветки, грозди, ягоды и семена. Почки у винограда могут осуществлять функции роста и плодоношения и давать начало вегетативным и одновременно генеративным органам.
Корень и корневая система.
Корни выполняют очень важную жизненную функцию растения. Они поглощают из почвы воду с растворенными в ней минеральными веществами, которые под давлением по проводящей системе передвигаются вверх в надземную часть к стеблям, листьям и генеративным органам. Молодые корни (мочки) наряду с растворами минеральных солей способны поглощать из почвы углекислый газ. Старые скелетные корни прочно закрепляют растение в почве. Они обычно длинные, мясистые, равномерно утолщенные, снаружи покрыты тонким слоем ежегодно отделяющейся корки. Скелетные корни служат не только проводниками воды с растворенными в ней питательными элементами по стеблю к листьям и генеративным органам. В них главным образом происходит синтез органических веществ, откладываются запасные питательные вещества (крахмал, белок, жиры), необходимые для растения в критические периоды его жизни. Исследованиями установлено, что при отсутствии пополнения запасов питательных веществ скелетные корни могут обеспечить жизнедеятельность куста до 6 лет.
Совокупность молодых и старых корней составляет корневую систему растения. Важная функция корней, особенно молодых,- превращение углеводов в аминокислоты и осуществление первичного синтеза белка. Образовавшиеся в листьях в процессе фотосинтеза углеводы быстро передвигаются вниз по стеблю и в корнях превращаются в сахара и различные органические кислоты, которые, взаимодействуя с аммонийными солями, образуют смеси аминокислот - основных компонентов белковых веществ. Незначительная часть органических кислот выделяется корнями в почву и вместе с разложившимися остатками корней обогащает ее органическими веществами. Это способствует развитию и скоплению в прикорневом слое почвы полезных микроорганизмов (грибы, бактерии и др.) в результате жизнедеятельности которых труднодоступные для растений питательные элементы почвы превращаются в легкоусвояемые формы. Из корней основная часть аминокислот поднимается вверх и концентрируется в тканях растущего побега и гроздей, используется для построения вновь образующихся клеток. Формирование и рост молодых побегов и почек зависят от прохождения всех этих процессов.
Корни винограда содержат 40-60% воды, по сравнению со стеблем в них больше азота, фосфора и зольных элементов, но меньше калия. Из пластических веществ в корнях больше всего углеводов, содержание которых заметно изменяется в течение сезона. В сентябре в корнях бывает 7% моносахаров, 3 - дисахаров и 13% крахмала. Наибольшее количество крахмала (23%) установлено в тонких корнях, наименьшее (15%) в толстых. Содержание крахмала увеличивается в ноябре. К. этому времени им заполнены коровая часть корня и сердцевинные лучи. В позднеосенний и зимний периоды до начала сокодвижения в корнях содержится 1,5-3% сахаров и свыше 20% крахмала. Кроме того, в зоне камбия корней имеются жиры, которые начинают накапливаться во второй половине лета (июле), а максимальное их количество наблюдается в зимний период.
Характер развития корней и корневой системы в целом зависит от способа размножения винограда, видовых и сортовых особенностей, физико-механических свойств, химического состава почвы, ее температуры, влажности и метода предпосадочной обработки.
При семенном размножении во время прорастания семени сначала через трещину его клювика из первичного корешка зародыша развивается главный, стержневой корень, на котором вскоре образуются корневые волоски. Через несколько дней главный корень начинает ветвиться, на его утолщениях появляются более тонкие боковые корни 1-го порядка, от которых отходят корни 2-го порядка, затем 3-го и т.д. От стержневого корня они идут сначала под прямым углом, а затем кончики их изгибаются и они растут почти параллельно главному корню. Стержневой корень сеянца растет сильнее боковых и в течение первого года может углубиться в почву до 1 м и более. В первые 2 года корневая система развивается значительно сильнее надземной части. У сеянца место перехода от корня к стеблю называют корневой шейкой.
При размножении винограда вегетативным способом - черенками, прививкой, отводками - при благоприятных условиях для их укоренения на узлах, реже на междоузлиях, корни развиваются из корневых зачатков клеток перицикла стебля против сердцевинных лучей. В этом случае образуются придаточные корни, которые называют также адвентивными. Достигнув длины 8-10 см, они начинают ветвиться, образуя боковые корни разных порядков, имеющие на концах утолщения (мочки) белого цвета. При посадке коротких одноглазковых черенков, которые размещают обычно в горизонтальном положении, так же как и отводков, корни развиваются пучком с противоположной от глазка стороны. При посадке длинных 3-, 4-, 5-глазковых черенков, которые размещают вертикально, корни развиваются на узлах черенка, сначала на нижних, а затем и на верхних, но больше всего на базальной части черенка - пятке.
Рис. 16. Корневая система:
/ - сеянца, // - саженца; 1 - стержневой корень; 2 - боковые корни; 3 - корневая шейка; 4 - поверхностные (росяные) корни; 5 - боковые корни; 6 - основные (главные) корни; 7 - подземный штамб.
В отличие от сеянца корневая система саженца (растения, выращенного из черенка) имеет мочковатый характер. Подземная часть саженца состоит из стебля (подземный штамб) с отходящими от него несколькими ярусами корней (рис. 16). В верхней части подземного штамба на глубине 10-15 см в ежегодно обрабатываемом верхнем слое почвы развивается большое количество поверхностных (росяных) корней, или росособирателей. Эти корни тонкие, короткие, и если их своевременно не удалить проведением катаровки, то растение полностью перейдет на развитие корневой системы в поверхностном слое почвы, что препятствует развитию корней в глубоких слоях, приводит к ослаблению кустов и их полной гибели. Это наблюдается при продолжительной засухе летом и после сильных морозов зимой. На привитых виноградниках поверхностные корни удаляют особенно тщательно, в противном случае кусты перейдут на корни привоя, и цель прививки как одного из способов защиты винограда от филлоксеры не будет достигнута. В средней части подземного штамба, преимущественно на месте бывших узлов, развиваются боковые корни, а у самого его основания - главные (пяточные), наиболее толстые, длинные, глубоко проникающие в почву. К трем-четырем годам у виноградного куста развивается мощная корневая система.

Рис 17. Анатомическое строение молодого корня в зоне поглощения (первичное строение):
1 - корневые волоски; 2 - эпидермис; 3 - интеркутис; 4 - эндодерма; 5 - первичная ксилема; 6 - первичная флоэма; 7 -кодовая паренхима; 8 - клетки, содержащие рафиды; 9 - перицикл.
Корень растет путем деления клеток образовательной ткани - меристемы, находящейся в концевой его части в конусе нарастания. Как у стержневых, так и у адвентивных корней кончик корня размером в несколько миллиметров снаружи покрыт твердым, острым и очень прочным чехликом желтого цвета. У воздушных корней, которые легко развиваются в условиях повышенной температуры и влажности воздуха, чехлик коричневый. Под чехликом находится зона роста. Это наиболее нежная часть корня длиной 2-5 мм, белого цвета. Клетки ее, вытягиваясь в длину, толкают вперед чехлик, под прикрытием которого верхушка корня прокладывает себе путь между частицами почвы. Этим объясняется способность винограда расти на каменистых, скелетных и других твердых почвах. Наружные клетки чехлика постепенно высыхают, отслаиваются и взамен их на кончике корня непрерывно образуются новые клетки из конуса нарастания. За зоной роста идет зона поглощения. Она более утолщена, желто-белого цвета, длиной 2-7 см. Наружные клетки ее (эпидермис) вытягиваются в длину, образуя множество корневых волосков. Оболочка корневого волоска не имеет кутикулы, поэтому поглощаемая ими из почвы вода с растворами солей легко проникает в клетки. Корневые волоски недолговечны, живут 10-20 дней. По мере роста и удлинения корня старые волоски отмирают, а взамен их образуются новые, и зона поглощения перемещается. За зоной поглощения следует проводящая зона. Снаружи эта зона покрыта толстым слоем пробки коричневого цвета, с возрастом корней она превращается в корку. В месте перехода зоны поглощения в проводящую зону у корня заметна тонкая перемычка. До нее корень считается молодым, а после нее - скелетным. По анатомическому строению молодой корень (рис. 17) значительно отличается от старого, скелетного. На поперечном срезе молодого корня, сделанном в зоне поглощения, снаружи видны сомкнутые однородные клетки эпидермиса с корневыми волосками. Под эпидермисом находится первый слой коры - интеркутис. За ним следуют 20-25 слоев клеток коровой паренхимы, а затем слой клеток с уплотненными стенками - эндодерма. В середине корня находится центральный цилиндр, который у молодого корня состоит из двух-трех первичных сосудисто-волокнистых пучков, расположенных по кругу.
Рис. 25. Переходные формы усика к соцветию:
а - внепазушный побег; б -переходы от внепазушного побега к усику; в- от усика
к соцветию.
Чашечка недоразвитая, состоит из 5 сросшихся недоразвитых чашелистиков, охватывает основание цветка слабовыраженными тупыми зубчиками. Венчик состоит из 5 сросшихся вверху лепестков, прикрывающих внутренние части цветка. Между чашечкой и основанием венчика находится небольшой кольцевой валик из недоразвитых нектарников, окрашенных в зависимости от сорта в зеленый, желтый либо в оранжевый цвет. В них содержатся эфирные масла с приятным запахом, напоминающим запах резеды, но. нектара они не выделяют, поэтому и не привлекают пчел. Тычинок в цветке 5, реже 6-7. Они расположены против лепестков венчика и слегка изогнуты. На конце тычиночных нитей находятся пыльники, содержащие пыльцу. Пыльники 2-гнездные, с широким связником, прикрепленным основанием к тычиночной нити. В каждом гнезде по 2 пыльцевых мешка, которые во время цветения открываются продольными щелями, в результате чего из них высыпается пыльца.
Пестик бутылкообразной или грушевидной формы, состоит из завязи, столбика и рыльца. Завязь верхняя, 2-гнездная, состоит из двух плодолистиков. В одном гнезде 2 анатропные семяпочки. В каждой из них находится многоклеточное ядро - нуцеллус с двумя покровами (интугементами) - внутренним и внешним, между которыми находится узкий канал - пыльцевход. В нуцеллусе семяпочки из археспоральнои клетки развивается зародышевый мешок, содержащий яйцеклетку, синергиды, полярные ядра и антиподы.
Наряду с общей схемой строения (2-гнездная завязь с двумя семяпочками в каждом гнезде) у многих сортов встречаются отклонения (1-8%), выражающиеся в наличии большого числа гнезд в завязи и семяпочек в них. Этим объясняется наличие более четырех семян в ягоде.
Столбик пестика цветка у разных видов и сортов винограда может быть коротким и толстым, удлиненным и тонким. Рыльце блюдцеобразной формы, с беловатыми приподнятыми ровными, реже вальчатыми краями.
У винограда несколько типов цветков (рис. 26): обоеполые (облигатно-фертильные); функционально-женские (факультативно-фертильные); истинно женские и мужские (облигатно-бесплодные).
У дикого винограда 2 основных типа цветка: мужской с различными вариациями и женский.
Большинство культивируемых сортов европейско-азиатского винограда имеют обоеполый тип цветка. Как исключение в старых насаждениях встречаются сорта с функционально-женским, в
насаждениях маточников филлоксероустойчивых подвоев - сорта в основном с мужским типом цветка. Все названные типы цветка различаются по морфологическим признакам. От умения распознавать тип цветка зависит правильное размещение сортов в насаждениях и организация работ по искусственному опылению самостерильных (бесплодных) сортов, урожай и качество винограда.
Рис. 26. Тип цветка винограда:
1 - обоеполый; 2- функционально-женский; 3 - истинно женский; 4 - мужской.
Цветок обоеполого типа имеет прямостоячие тычинки, которые выше пестика или равны ему. В пыльниках находится жизнеспособная (фертильная) пыльца. Завязь хорошо развита, обычно грушевидной формы. Пыльца обоеполого цветка в сухом виде имеет правильную удлиненно-бочкообразную форму, в случае погружения в 10- 15%-ный раствор сахара (глюкозы) при температуре 25-30°С легко прорастает в пыльцевую трубку. Сорта винограда с обоеполым типом цветка в основном самоопыляющиеся (автогамия), но встречаются сорта, нуждающиеся в перекрестном (аллогамия) опылении.
Цветок функционально-женского типа также имеет пестик и тычинки, но по морфологическому строению отличается от обоеполого. Тычинки менее развиты, короче пестика, пестик довольно развитый. Во время цветения после сбрасывания колпачка тычинки отгибаются книзу.
Пыльца бесплодная - стерильная, в сухом виде имеет неправильную форму: угловатую, ромбическую, трехгранную, округлую, овальную, шаровидную и т.д. В растворе сахара пыльца цветка этого типа не прорастает в пыльцевую трубку, а только едва набухает и принимает округлую форму. Сорта с таким типом цветка самостерильны и нуждаются в перекрестном опылении. Бесплодность пыльцы и недостаточное искусственное опыление у сортов с функционально-женским типом цветка - причины чрезмерного осыпания цветков, а иногда и засыхания соцветий. Если сорта винограда с функционально-женским типом цветка своевременно не опылить, то ягоды на них или не завязываются, или бывают мелкими, величиной с горошину (партенокарпические), без семян либо с мелкими Щуплыми семенами без зародыша и эндосперма.
Мужские цветки винограда имеют сильноразвитые прямостоячие тычинки с большим количеством жизнеспособной пыльцы в пыльниках, завязь недоразвита, рыльце на пестике отсутствует, сам пестик имеет различную форму, от маленького бугорка до сильноразвитой крупной завязи. Пыльца цветка мужского типа, как и у обоеполого, в сухом виде имеет правильную удлиненно-бочкообразную форму. Она также хорошо прорастает в растворе сахара в течение нескольких часов в пыльцевую трубку. Растения с мужским типом цветка имеют крупные соцветия, но ягод
не образуют. В том случае, если по внешним морфологическим признакам трудно определить тип цветка, его устанавливают по форме и способности прорастания сухой пыльцы.
Соцветия с мужским типом цветка используют для сбора пыльцы, необходимой для опыления сортов винограда с функционально-женским типом цветка. Есть и очень ценные старые сорта, такие как Нимранг, Чауш белый, Пухляковский, Тавквери, Катта-Курган и другие, с функционально-женским типом цветка, нуждающиеся в перекрестном опылении. В посадках винограда к таким сортам подбирают соответствующие опылители, у которых совпадает срок цветения. Их высаживают рядами - 1 ряд сорта-опылителя и 2- опыляемых. Кроме того, применяют искусственное опыление. Однако из-за технологических сложностей возделывания и снижения урожайности в годы с неблагоприятными для опыления погодными условиями новые сорта с таким типом цветка Государственной комиссией по сортоиспытанию сельскохозяйственных культур Госагропрома СССР в настоящее время не принимаются.
Помимо основных указанных типов цветка, у винограда встречаются и истинно женские цветки (клон сорта Мурведр), а также различные аномалии в строении цветка. Чаще всего аномалии проявляются в раскрывании цветка розеткой, звездочкой или в махровости. Махровость присуща в основном сортам, имеющим функционально-женский тип цветка. Встречаются и фасциации цветков, выражающиеся в слиянии двух цветков в один сложный или в появлении цветков со сросшимися двумя-тремя цветоножками в одну и др.
Гроздь, ягода, семя.
Соцветие после оплодотворения и естественного опадения цветков и завязей растет и формируется в гроздь. Ножка соцветия преобразуется в ножку грозди, ось соцветия с разветвлениями - в гребень, завязи - в ягоды. В процессе роста и развития гребня в нем наиболее сильно развиваются механические и проводящие ткани, в частности мягкий луб, что обеспечивает прочность гребня и приток к ягодам большого количества пластических веществ. Ножка грозди, при помощи которой гроздь прикрепляется к побегу, в зависимости от сорта бывает короткой и длинной. У одних сортов ножка все время остается травянистой, зеленой, а у других она древеснеет. На ножке, недалеко от места ее прикрепления к побегу, имеется утолщение - узел, от которого ответвляется усик. Если усик имел на себе цветки, то после их опыления усик превращается в ответвление основной грозди, называемой крылом. У некоторых сортов их может быть 1-2. Крылья отличаются от основной грозди меньшей величиной и меньшим количеством ягод. Иногда крылья развиваются наравне с основной осью грозди.

Рис 27. Строение ягоды винограда, продольный разрез:
1 . плодоножка; 2 - подушечка; 3 - сосудистые пучки; 4 - семена 5 - кожица;
6 -
мякоть; 7 - сердечко; 8 - эпидермис.
Разветвленная часть гребня заканчивается плодоножками, на которых сидят ягоды. От характера ветвления гребней и длины осей 1-, 2-го и последующих порядков зависят величина, форма и плотность грозди, которые являются сортовыми признаками и имеют важное практическое значение. Эти показатели могут изменяться в зависимости от почвенно-климатических условий и приемов агротехники. Однако грозди, расположенные ближе к основанию плодоносного побега, всегда крупнее вышерасположенных.
На рост гроздей и ягод винограда значительно влияет обработка гиббереллином и другими регуляторами роста.
Величина грозди определяет урожайность сорта и удобство ручной уборки. Чем больше средняя масса грозди, тем выше урожай с 1 га насаждений.
Анатомическое строение ножки грозди и плодоножки ягоды аналогично строению междоузлия молодого побега.
Ягода - основной орган, ради которого культивируют виноград. Она развивается из завязи цветка. После оплодотворения яйцеклетки образуется зародыш, который стимулирует рост завязи (околоплодника). Рыльце и пестик высыхают, превращаясь в пупок, хорошо заметный на вершине вызревшей ягоды. К гребню грозди ягода прикреплена с помощью плодоножки, расширенной у основания в подушечку (рис. 27). Через плодоножку и подушечку из гребня в ягоду проходят сосудисто-волокнистые пучки, которые в виде мелкой сетки расходятся под кожицей мякоти ягоды и идут к семенам. При отрыве ягод от плодоножки на подушечке остается кисточка. Это оторванные сосудисто-волокнистые пучки.
Кожица ягод (эпикарпий) состоит из однослойного эпидермиса толщиной 30-40 мкм, клетки которого напоминают правильный многоугольник, и 10-15 слоев клеток кожицы, расположенных под эпидермисом. В эпидермисе имеются щели - устьица, через которые происходит газообмен. Сверху клетки эпидермиса утолщены и кутикулонизированы. Количество слоев кожицы, их толщина и плотность обусловливают устойчивость ягод к загниванию, а у столовых сортов - транспортабельность и пригодность для длительного хранения. С ростом ягоды клетки кожицы растягиваются, становятся округлыми и переходят в радиально вытянутые клетки мякоти - мезокарпий. Это основная часть ягоды клетки мякоти не прилегают друг к другу. Между ними имеются большие сообщающиеся между собой межклеточные пространства, через которые осуществляется газообмен. В мякоти выделяются 2 зоны. Первая зона находится непосредственно под кожицей и состоит из нескольких слоев крупных радиально распложенных клеток, достигающих 0,3-0,4 мм в диаметре. Вторая зона - это самый внутренний, прилегающий к семенам слой (эндокарпий, сердечко), состоящий из 5-6 слоев удлиненных клеток. Общее число слоев клеток ягоды от эпидермиса до эндокарпия такое же, как у завязи (около 25-30). У столовых сортов винограда количество слоев клеток кожицы и сами клетки больше, ко времени созревания они сильно растягиваются. Клетки мезокарпия, прилегающего к кожице, имеют плотную мембрану и выделяют небольшое количество сока, что обусловливает их характерную мясистую, мясисто-сочную, плотную хрящеватую, хрустящую консистенцию.
У столовых сортов кожица не отделяется от мякоти и при нагрузке раздавливается вместе с ней. У технических сортов число слоев клеток кожицы меньше и сами клетки мельче. Мезокарпий же состоит из более крупных клеток с очень нежными мембранами, которые при созревании винограда растворяются, вследствие чего мякоть ягод представляет сплошную жидкую массу. Кожица у большинства винных сортов тонкая, эластичная, легко разрывающаяся, с большим количеством сока в вакуолях, легко отделяется от мякоти при раздавливании.
Оболочка клеток мякоти, представленная клетчаткой и целлюлозой, составляет 0,3-0,5% массы мякоти, остальное - это клеточный сок, содержащий у зрелых ягод до 55-95% воды, 5-30- сахаров (глюкоза, фруктоза, сахароза), 0,5-1,9% органических кислот (винная, яблочная, глюконовая, глюкуроновая, лимонная и др.). Витамины (А, В1 В2, РР, С) содержатся в основном в кожице. В мякоти и соке их меньше. Дубильные вещества, так же как крахмал, имеются в мякоти только до начала созревания ягод, затем их количество резко уменьшается и ко времени созревания ягод они полностью исчезают. Кроме того, в ягодах накапливаются ароматические вещества, которые представляют собой сложный комплекс летучих ароматических эфирных масел, обусловливающих аромат ягод. Они накапливаются вблизи проводящих пучков, примыкающих к кожице.
Пока ягода растет, она дышит, ассимилирует и остается зеленой. В ней содержатся хлорофилл и красящие вещества - каротин, ксантофилл и др. Красящие вещества накапливаются в ягоде постепенно. Максимальное их количество наблюдается в период полной зрелости ягод. В процессе созревания ягод вещества, обусловливающие их первоначальную окраску, разрушаются, и в клетках кожицы, в одном, двух или во всех ее слоях, накапливаются другие пигменты - флавонолы, основные компоненты которых- кверцитин и кверцинтрин, обладающие желтой окраской. Это продукты изменения хлорофилла, каротина, ксантофилла и других сопутствующих хлорофиллу веществ. Они придают сортам с белой ягодой окраску от светло-зеленой до золотисто-янтарной. Другие пигменты, представляющие антоциановый комплекс, в состав которого входят ампелопсин, энин и прочие, обусловливают образование красных веществ, придающих ягодам розовую, красную и черно-синюю окраску. Цвет ягод - четкий ампелографический признак. Однако в зависимости от экологических условий выращивания винограда у одного и того же сорта он может изменяться: быть светлее или темнее. Так, например, у сортов Тайфи розовый, Октябрьский, Забалканский в условиях Крыма ягоды приобретают интенсивно-розовую и даже красную окраску, тогда как в Средней Азии ягоды у этих сортов светло-розовые. В клетках мякоти красящих веществ обычно не бывает, но есть сорта (Пти Буше, ВИР 1, Одесский черный, Саперави и др.), красящие пигменты у которых находятся и в вакуолях клеток мякоти. Это так называемые сорта-красильщики, из которых можно приготовить только интенсивно окрашенные соки и вина.
На поверхности кожицы, особенно у сортов со светлой окраской ягод, заметны коричневые точки - чечевички, или остатки опробковевших ко времени созревания ягод устьиц. Снаружи кожица ягод покрыта восковидным сизо-голубоватым налетом - пруином, наиболее ярко выраженным у темноокрашенных сортов. Пруин выполняет защитную роль: предохраняет ягоду от неблагоприятных внешних условий, потери воды, гниения. Ко времени созревания кожица ягод в зависимости от сорта становится тонкой, легко рвущейся, прозрачной, не ощущаемой при еде или толстой, грубой и очень плотной. Ягоды разных сортов винограда различаются по величине, форме, окраске и вкусовым качествам. Мякоть ягоды может быть сочной, тающей, плотной, хрустящей, мясистой, слизистой. У сортов винограда с сочной мякотью ягод стенки клеток тонкие, легко разрываются при раздавливании, и сок при этом быстро вытекает. Сорта с такой ягодой нетранспортабельны и не хранятся. Наоборот, у сортов с плотной хрящевой мякотью ягоды менее сочные и при раздавливании плохо отдают сок. Такие сорта пригодны для длительного хранения и транспортировки на дальние расстояния. Вкус ягод определяется гармоничным сочетанием в мякоти сахара и кислоты. Ягоды многих сортов винограда имеют специфический аромат (группа мускатных сортов - Мускат белый, Мускат черный, Мускат венгерский, а также сорта Рислинг, Траминер розовый, Каберне Совиньон, Изабелла, Лидия).
Некоторые сорта (группы Шасла, Португизер и др.) выраженного аромата не имеют.
Величина (масса) ягоды зависит от числа семян в ней. С увеличением числа семян от одного до четырех увеличивается масса ягоды. На каждое семя приходится около 10% массы ягоды. От того, как прошло оплодотворение и сколько развилось семян в ягоде, зависит масса ягоды и всей грозди, а следовательно, и величина урожая с куста и единицы площади.
Ягоды у винограда бывают с семенами и без семян. Бессемянных сортов немного. Из общего числа районированных в СССР сортов винограда (свыше 250) бессемянных только 9, или около 4%.
Согласно схеме происхождения бессемянных сортов винограда, разработанной и предложенной К. В. Смирновым, первые бессемянные формы винограда появились в виде мутаций на семенных сортах. Затем в результате естественной гибридизации между семенными и бессемянными сортами возникли вторичные бессемянные формы. Путем целенаправленной селекции сортимент их был расширен. Бессемянные сорта делят на 2 группы: коринок (белая, розовая, черная), у которых образование и рост ягод проходят за счет разрастания завязи до величины горошины, без процессов опыления и оплодотворения (партенокарпия); кишмишей (белый, розовый, черный и новые бессемянные сорта), которым для образования ягод необходим процесс опыления. Процесс же оплодотворения нарушается на различных этапах развития, что приводит к образованию мелких недоразвитых семян (рудиментов), значительно различающихся по величине и степени развития (стеноспермокарпия). У отдельных бессемянных сортов и гибридных форм может встретиться 2 формы бессемянности - партенокарпия и стеноспермокарпия. Так, у клонов Кишмиша белого в одной грозди можно одновременно обнаружить ягоды бессемянные, овальной формы, свойственной сорту величины (кишмишность) и круглые, мелкие (партенокарпия). Имеет место и частичная партенокарпия, когда в нормально развитой грозди с развитыми семенами в ягодах часть ягод остаются бессемянными, недоразвитыми (горошащимися), что наблюдается при плохом опылении сортов с функционально-женским типом цветка (Мадлен Анжевин, Чауш белый и др.). В этом случае в процессе оплодотворения пыльцевые трубки достигают нуцеллуса, но не входят в него, а переплетаются между собой, образуя плотный клубок, и в таком виде отмирают.
Семена у винограда мелкие, грушевидной формы, с заостренным удлиненным клювиком. Пока ягода растет, семена находятся в молочной зрелости, мягкие по консистенции и зеленые по цвету. Ко времени созревания семена покрываются прочной кожурой, содержащей каменистые клетки, богатые фенольными веществами. Благодаря этому они становятся твердыми и приобретают желтовато-коричневую окраску. Под кожурой семени находится эндосперм, клетки которого богаты питательными веществами - белком, жиром. В клювике семени расположен зародыш, несущий зачатки органов будущего растения: корешок, обращенный к микропиле, стебелек, 2 семядоли сердцевидной формы, между которыми расположена зародышевая верхушечная почечка - эпикотиль. Семя винограда имеет брюшную и спинную сторону. На брюшной стороне 2 продольные бороздки - впадины, между которыми проходит семенной шов, переходящий на более выпуклую округлую спинную сторону до халазы - места проникновения в семя сосудисто-волокнистых пучков. В ягодах содержится обычно 2-3 семени, но бывает 4 и более. Как указывалось выше, есть и бессемянные сорта винограда, которые, согласно классификации К. В. Смирнова, по величине рудиментов семян и степени их развития делят на 3 категории.
К первой категории относят сорта винограда, в ягодах которых содержатся мелкие (до 6 мг) рудименты семян (Кишмиш белый круглый, Кишмиш белый овальный, Кишмиш розовый, Кишмиш черный), ко второй - сорта с рудиментами семян 6,1 -10 мг (Аскери, Бедона), к третьей категории - с более крупными рудиментами- 10,1 - 14 мг (Кишмиш люнда и др.).
Семена разных видов и сортов винограда различаются по величине, форме, окраске, длине клювика, расположению и форме халазы. У сортов V . vinifera
семена крупные и имеют удлиненный клювик. Халаза расположена в верхней трети спинной стороны, тогда как у других видов - в ее средней части. Кожура семени состоит из трех, слоев - наружного, промежуточного и внутреннего. Наружный слой имеет один ряд тангентально удлиненных клеток, промежуточный - несколько рядов тонкостенных клеток рыхлого строения, служащих у невызревших семян вместилищем крахмала и рафид, а у вызревших они высыхают, сплющиваются и становятся местом накопления танина. Внутренний, защитный слой примыкает к эндосперму и состоит из двух - шести рядов радиально удлиненных каменистых клеток с одревесневшими стенками. Оболочка из каменистых клеток полностью окружает семена винограда, кроме халазы и верхушки клювика, где находится отверстие - микропиле. В клювике каменистых клеток значительно больше, чем в остальной части семени.
Шаламов В.Н. | 2015-02-28
По данным исследований, в условиях средней зоны за вегетационный период у однолетнего сеянца яблони китайской образуется до 40 тысяч корней, имеющих суммарную длину до 230 метров. Однолетние сеянцы яблони лесной и сибирской, груши дикой, вишни и других видов плодовых растений развивали за это время гораздо меньше корней (по длине и количеству), по сравнению с сеянцами яблони китайской.
Длина всех корней взрослого плодового дерева выражается десятками километров, а суммарное количество - миллионами отдельных корешков. Подавляющая часть корней имеет малую длину. Например, по результатам одного из исследований, у яблони корни длиной от долей миллиметра до 5 мм составляют 65,1%, у груши - 43%; корни длиной от 6 до 10 мм - соответственно 18,7 и 23%.
Остальные корни имеют длину от 1–2 см до 0,5 м, но их гораздо меньше (у яблони 16,2%). Средняя длина корня у однолетних сеянцев плодовых растений довольно постоянна: у яблони - 6–7 мм, у груши - 8–9, у вишни - 8 мм. У плодоносящих деревьев плодовых растений средняя длина корня в 2 раза меньше (у яблони 3,5 мм), чем у однолетних сеянцев.
В процессе жизни растения у его корневой системы происходит циклическая смена (самоизреживание) отдельных корней. Проведенные исследования позволили установить, что у каждого растения с первых дней и до конца жизни происходит неуклонное и последовательное отмирание концов осевых корней, а также коротких боковых корней сначала на главном корне, затем по мере роста на более длинных корнях последующих порядков ветвления (первом, втором и так далее). Далее наблюдается отмирание целых мочек (сеток) корней и замена их новыми, то есть очищение (оголение) верхних, уже более толстых корней от отдельных более мелких корней и мочек.
На однолетних сеянцах за один сезон могут отмирать десятки тысяч, а у взрослых деревьев - миллионы корней. Это свидетельствует о недолговечности всасывающих корней, которые в зависимости от расположения в корневой системе живут от нескольких дней до месяца и редко дольше.
Отмирание корней (корнепад)
Отмирание корней было названо «корнепадом», одним из ведущих исследователей корневой системы плодовых и ягодных растений, в период СССР, Колесниковым В. А. Это название достаточно прижилось и используется и сейчас. Оно несколько созвучно с явлением листопада у плодовых, ягодных и орехоплодных растений.
У разных видов растений отмирание корней может ускоряться или замедляться в разной степени, в зависимости от условий жизни. В результате отмирания и возобновления корневая система растения непрерывно в течение вегетации и всей жизни продвигается во все новые, неиспользованные слои почвы. Она распространяется от ствола круговыми полосами, отмирая в одном месте и появляясь в другом, и обеспечивает нормальные условия роста и плодоношения растений.
Суммарная масса отмерших в течение года корней выражается десятками килограммов и даже тоннами органических остатков в расчете на гектар, являющихся источниками питания микроорганизмов ризосферы.
В процессе корнепада происходит отмирание не только обрастающих (мелких корней), но и скелетных. Отмирание и возобновление корней, или циклическая смена их, у сеянцев и взрослых плодовых, ягодных и орехоплодных растений представляет естественный процесс в жизненном цикле их развития и свойственно всем древесным и травянистым растениям.
Зависимость от происхождения

Рост и формирование корневой системы в почве зависит от происхождения корней (семенные или вегетативные), вида и сорта растения (привой), природных условий и агротехники. Корневые системы семенного происхождения обычно проникают в почву глубже, чем вегетативного происхождения. Различны типы роста корней в стороны и в глубь почвы, а также их расположение по горизонтали почвы и почвогрунта.
Например, в средней полосе ежегодный прирост корней плодовых растений в сторону от ствола по радиусу составлял около 20–36 см, а в глубину - 17–33 см. Различия значительны, и садовод должен учитывать их при выборе и подготовке почвы под сад и при дальнейшем уходе за корнями и почвой.
Агротехническими приемами, например плантажной обработкой и удобрением, на любой почве, в том числе и на распространенной у нас дерново-подзолистой, можно увеличить массу корней, расположенных ниже пахотного горизонта. Тонкие, обрастающие и всасывающие корни в зависимости от природных условий и агротехники могут расти с ранней весны и до поздней осени, даже после листопада, а затем прекращать рост и отмирать.
В общем, постепенно разрастаясь, корневая система сеянца или саженца, а позднее взрослого плодового, ягодного или орехоплодного растения охватывает огромные объемы почвы и почвогрунта длинными и толстыми тяжами корней, на которых размещены сотни тысяч коротких корешков, а на них десятки миллионов коротких волосков.
Корни горизонтального направления у плодовых пород в северной зоне в массе располагаются на глубине примерно до 30–50 см, в средней - до 50–75 см, у большинства ягодных пород в этих же зонах - на глубине 0–60 см, у карликовых деревьев яблони - на глубине 0–50 см, у стланцевых деревьев яблони и груши - на глубине 0–40 см.
В результате наличия плотных, а местами сцементированных прослоек и погребенных горизонтов в почве корневая система может залегать ярусами. Например, в Среднем Поволжье на дерново-подзолистой почве первый ярус корней яблони залегает на глубине 18 см, второй - 101–140 см. Более тонкие корни в значительном количестве могут встречаться во всех зонах садоводства в поверхностном слое почвы с глубины 3–10 см. У яблони в северной зоне корни чаще размещены глубже 10–20 см от поверхности почвы.
Проведенные исследования показали, что у плодовых растений, начиная со второго года и всю последующую жизнь, диаметр корневой системы в 1,5–2 раза больше диаметра (проекции) кроны. Такое соотношение диаметра крон и корневых систем наблюдается у подавляющего большинства плодовых, ягодных и орехоплодных растений во всех зонах садоводства.
Опыты, проведенные в средней полосе, показали, что у 7-летних деревьев яблони корневая система разрастается в стороны по диаметру на 3,5 м, 14-летних - на 5 м, у 20-летних - на 8–9 м. При этом выявился следующий порядок распределения разных видов плодовых растений по диаметру распространения корневой системы - абрикос, яблоня, груша, слива, вишня.
Корни вертикального направления у разных плодовых, ягодных и орехоплодных растений проникают в северной зоне в большинстве случаев до 1–2 м, в средней - до 2–4 м, а в южной - до 5–10 м. Вглубь корни продвигаются главным образом по ходам землероев, особенно дождевых червей, и по трещинам почвы. На глубину проникновения корней сильное влияние оказывают почвенные условия, привой и подвой.
Так, на дерново-подзолистой почве в Московской области корни яблони проникают на глубину до 3–4 м, груши, сливы и вишни - до 2 м. На этой же почве основная масса вертикальных корней Антоновки обыкновенной на китайке залегает на глубине 1 м, на полукарликовом клоновом подвое - 1,3 м, на яблоне лесной и сеянцах Аниса - 2 м, а на сеянцах Антоновки - на глубине 2,5 м. Количество и длина корней вертикального направления может быть значительным, особенно у плодовых деревьев, выращиваемых в южной зоне. В этой зоне корни даже карликовых клоновых подвоев могут достигать глубины 5 м.
Как правило, чем южнее произрастают садовые растения, тем глубже залегают их корневые системы. Однако и на юге корневые системы могут быть расположены поверхностно. Это наблюдается в случаях близкого залегания грунтовых вод или плотных, а иногда галечных прослоек и сцементированных отложений. В таких случаях данные растения оказываются менее урожайными и менее долговечными.
Например, в Крыму деревья яблони сорта Сары Синап в возрасте 35 лет при размещении корней на глубине до 4,6 м давали до 300 кг плодов, а деревья того же сорта и возраста при других почвенных условиях (близкие грунтовые воды) и глубине залегания корней до 1,5 м давали только 100 кг.
Глубина залегания корней растений
Глубина проникновения корней в почву имеет очень большое значение. Специалисты указывают, что деревья яблони в средней зоне, имеющие глубину залегания корней до 3 м и больше, гораздо лучше развивались и плодоносили, чем деревья с глубиной проникновения корней до 1,6 м, и что деревья яблони с поверхностными корнями (мочкой) больше нуждаются в удобрении и болезненно реагируют на всякие недочеты в агротехнике во время вегетации. Кроме того, крупные корни при поверхностном их залегании будут ежегодно повреждаться при обработке почвы, тонкие же корни намного хуже будут обеспечены в верхнем слое почвы влагой, что может привести к их отмиранию при отсутствии регулярных поливов. С более глубоким залеганием корневой системы связан и меньший выпад плодовых деревьев. Так, по наблюдениям в яблоневом саду в средней зоне при паровой обработке почвы, когда у деревьев была глубокая корневая система, общий выпад деревьев за 13 лет после посадки составлял 2,4% против 11,1% у деревьев с менее глубоко залегающей корневой системой при задернении почвы в саду.
Лучшей корневой системой садовых растений следует считать такую, которая залегает равномерно по кругу, более глубоко и широко и имеет возможность извлекать воду и питательные вещества из большого объема почвы и почвогрунта. Все указанные растения в этом случае характеризуются большей засухо- и морозоустойчивостью.
Исследованиями отечественными и зарубежными учеными установлено, что данные растения, особенно плодовые, с мощной корневой системой гораздо дольше живут, более зимостойки, лучше и регулярнее плодоносят. Поэтому очень важно соответствующими агротехническими приемами способствовать глубокому залеганию корневой системы. Вместе с тем садовод всегда перед началом обработки почвы должен проверять глубину залегания корней, чтобы правильно назначить на каждом садовом участке под каждым деревом и кустом глубину обработки, внесения удобрений и тому подобное.
При выращивании в саду плодовых, ягодных и орехоплодных растений наблюдается взаимовлияние их корневых систем. Особенно такое влияние (конкуренция) корней наблюдалась между разными экземплярами растений одного и того же вида. При этом корни растений распространяются в направлении слабого соседнего растения. При наблюдении за корневыми системами разных плодовых растений было отмечено:
- Корни яблони в одних местах не входят, а в других входят в корни соседней яблони.
- Корни яблони входят в корневую систему выкорчеванного в прошлом году дерева яблони, но уходят от корневых систем сеянцев яблони только что выкорчеванного питомника.
- Корни сливы, черешни свободно входят в корневую систему яблони, То же самое наблюдалось в отношении корней яблони, груши и абрикоса, миндаля, то есть между корнями этих пород нет конкуренции ни за пищу, ни за воду и они не испытывают никакого неудобства из-за присутствия корней растений других видов.
Проведенные исследования показали, что главную роль в совместимости корневых систем растений одного вида или разных видов играет аллелопатическое их действие посредством выделения в почву токсических веществ (колинов). Проявление корневыми системами плодовых, ягодных и орехоплодных растений явления аллелопатии следует обязательно учитывать при их выращивании.
Связь между ростом корневой системы и лиственным аппаратом

Научные наблюдения последних лет показали, что существует довольно тесная взаимосвязь между ростом и развитием корневой системы плодово-ягодных растений и ростом и развитие лиственного аппарата и надземной части растения вообще.
Например, выяснилось, что присутствие растущих корешков на корнях саженцев этих растений значительно сказывается на приросте их побегов. При наличии растущих корешков рост побегов после посадки саженцев яблони в сад был намного сильнее - 4 метра на одно растение против 1,5 метров у обычных растений.
Научное исследование суточного прироста корней и плодов у яблони сорта Славянка показало наличие определенной взаимосвязи - при усилении роста корней ослабевал рост плодов, а затем происходило обратное.
Также проводилось изучение соотношения размеров ветвей и корней, в процессе роста плодовых, ягодных и орехоплодных растений. Эти исследования показали, что, например, у молодых растений яблони, груши, сливы и других прирост корней неизменно превышал прирост ветвей, причем весьма значительно. Далее, наоборот, более усиленно происходил рост надземных органов.
Ранее агрономы предполагали наличие автономной связи между отдельными крупными скелетными корнями и отдельными скелетными ветвями. Однако, начиная с конца 50-х годов прошлого века, благодаря использованию для изучения процессов питания растений метода меченых атомов, были получены данные, которые не полностью согласуются с этим общепринятыми когда-то взглядами.
Например, в одном из опытов автономная связь между корнями и ветвями наблюдалась только у корнесобственных яблонь, но у привитых яблонь такой автономной связи не наблюдалось. В другом опыте указывалось, что наряду с локализацией основного количества радиофосфора в одной из скелетных ветвей кроны наблюдалось распространение небольших его количеств и в остальной части кроны.
Значение листового аппарата
Для роста корней очень большое значение имеет здоровый листовой аппарат. Так, в исследованиях летнее удаление боковых побегов приводило к задержке роста корней яблони.
Корневая система начинает хуже развиваться при нарушении нормальной деятельности листового аппарата, что бывает, например, при неправильной обрезке растений, повреждении листьев вредителями и болезнями, преждевременном листопаде и так далее.
Рост корней у плодовых деревьев с урожаем бывает более слабый, чем у деревьев без урожая. Отличается и сам характер роста корней - у деревьев с урожаем наблюдается медленный рост, быстрое побурение многих боковых разветвлений, более коротких, чем у деревьев без урожая.
Отставание роста корней на деревьях с урожаем обусловлено большим расходом питательных веществ на рост плодов, то есть тем же, чем объясняется слабый прирост побегов в урожайные годы.
Улучшить рост активных корней в урожайные годы и вместе с тем ослабить склонность растений к периодичности плодоношения можно хорошо известными способами - внесением удобрений, поливом, прореживанием цветков и надлежащей обрезкой.
Рост и развитие корневой системы
Были проведены и исследования по развитию корневой системы яблони в зависимости от развития надземной части. Оказалось, что существует определенная взаимосвязь между развитием всасывающей поверхности корня сеянца яблони и развитием листового аппарата. Отношение всасывающей поверхности корня к площадям листового аппарата оставалось почти постоянным в течение всей вегетации до периода листопада и равнялось 1,5–2.
На рост корневой системы очень сильное влияние оказывают внешние условия, а также приемы агротехники, в первую очередь связанные с воздействием на почву (обработка почвы, внесение удобрений, полив, борьба с болезнями и вредителями и так далее). Особенно большое влияние оказывает на развитие корневой системы аэрация почвы, водный, температурный и пищевой режимы почвы.
Аэрация корней растений

Значение аэрации для роста корней было показано во многих исследованиях. Эти исследования показали, что если около дерева или куста почва была разной плотности, то корни развивались в сторону рыхлой почвы.
На плотных почвах наблюдалось и более поверхностное развитие корневой системы. Лучшая аэрация давала эффект в виде большой, разветвленной, корневой системы. В одном из исследований в условиях вегетационного опыта при недостатке кислорода резко ослаблялось развитие и корневой системы, и надземных органов (появление признаков хлороза, усыхание листьев и другое).
Корневые волоски реагировали на аэрацию корневой системы моментально. Минимальным содержанием кислорода в данном опыте для корневых волосков яблони является 7,5 мл в литре воды, а для смородины - 5,8 мл.
Аэрация корней имеет важнейшее значение нормального развития растения и для его нормального питания. При недостатке в почвенном воздухе кислорода происходило нарушение поступления минеральных веществ в сеянцы яблони.
Другой вегетационный опыт показал, что однолетки яблони, посаженные в сосуды, набитые почвой разной степени уплотнения, а также почвой, смешанной с песком, давали разное количество новообразовавшихся корней, которое было значительно больше в рыхлой почве, а также в почве, смешанной с песком, и значительно меньшим в уплотненной почве.
Научные исследования показали, что корни свободно проникают в почву с плотностью ниже 30 кг на 1 см 2 , гораздо хуже при плотности 30–60 кг на 1 см 2 и совсем не растут при плотности выше 60 кг на 1 см 2 . Совершенно непроницаемыми для корней плодовых деревьев являются твердые прослойки земли с плотностью 300 кг на 1 см 2 .
Влияние воздуха на корневую систему растений
Огромный интерес представляет исследование влияния, на корневую систему растений, составных частей почвенного воздуха - кислорода, углекислоты и азота:
- Дыхание корней яблони ослаблялось при уменьшении содержания кислорода в почвенном воздухе ниже 8–9%, а также при повышении содержания угольной кислоты выше 5–6%.
- Недостаток кислорода особенно сказывался на состоянии корневых волосков: достаточно было 30-минутного пребывания всасывающего корня в бескислородной среде, чтобы корневые волоски не образовывались.
- Плодовые, ягодные и орехоплодные растения особенно чувствительны к недостатку кислорода в периоды сильного роста активных корней (весной и осенью).
- Больше кислорода требовалось для развития кончиков корней (5–10%) и еще больше для появления новых корней (12%).
- При содержании кислорода менее 10% снижался сухой вес корней, а ниже 15% - ослаблялось поглощение и накопление зольных элементов в растении.
- У большинства растений замена кислорода, в почвенном воздухе, азотом приводит к уменьшению всасывающей способности корней, а в дальнейшем - к их отмиранию.
Таким образом, для корней плодовых, ягодных и орехоплодных растений в почвенном воздухе обязательно необходимо присутствие кислорода, в то же время избыток углекислоты вредит им больше, чем недостаток кислорода.
При нормальном газообмене между почвой и атмосферой почвенный воздух наполняется кислородом и освобождается от избытка угольной кислоты, то есть процесс идет именно так, как это необходимо для лучшего развития корней.
Влияние влажности почвы на рост корней

Большую роль в росте корневой системы играет влажность почвы. Достаточная влажность почвы является особенно важной для активных мочковатых корней, которые развиваются лишь в достаточно увлажненных слоях почвы.
На сухих, малоорошаемых, почвах корни растений развиваются преимущественно в горизонтальном направлении, чем на почвах достаточно увлажненных. Поверхностное залегание корней наблюдается и при дерновой системе содержания почвы, например в яблоневом саду, по сравнению с паром. Очень большое значение для роста корневой системы имеет полив.
Так, в одном из опытов без орошения длина скелетных корней шестилетнего дерева яблони составляла 1558 м, то при орошении она равнялась 3219 м. Под влиянием орошения увеличилась также глубина залегания и распространения корней в горизонтальном направлении.
Другой опыт, с корневой системой яблони, показал, что она намного лучше развивалась, когда поливы делались при более высокой влажности, то есть когда поливами поддерживалась в течение всего вегетационного периода большая влажность почвы.
Отрицательное влияние оказывает на рост корней и избыток влаги, в связи с ухудшением условий аэрации. Этим, кстати, объясняется угнетающее влияние на корневую систему неглубокого залегания грунтовых вод.
Ослабление роста корней начинается задолго до начала увядания растений. Одно из исследований показало, что рост корней и надземных частей начал замедляться при постепенном нарастании засушливых условий за 7–11 суток до начала увядания листьев.
Поглощающие корни чувствительнее к недостатку влаги, чем проводящие - при длительной засухе они прекращали рост за 1–3 суток до начала увядания листьев. Прекращение роста этих корней сопровождается быстрой их субернизацией, и вслед за этим или одновременно начинается увядание листьев.
Отрицательно реагируют поглощающие корни и на воздушную засуху. В этом же исследовании рост их прекращался даже при кратковременном снижении влажности воздуха с 90–80 до 60–50%. Однако, хотя рост активных корней при недостатке влаги и прекращается, они могут до 3–4 месяцев оставаться живыми, не теряя способности частичного всасывания, и при благоприятных условиях восстанавливают свою активность.
Интересные данные были получены в исследовании о возможности передвижении влаги в самом растении в те участки корневой системы, которые испытывают недостаток в ней. Обнаружилось, что корни, находящиеся в сухой почве, могут частично получать влагу от корней, находящихся в более увлажненных слоях почвы.
Последним можно объяснить эффективность чересполосного задернения междурядий, хороший рост и высокую урожайность плодовых деревьев при глубоком залегании корней. Глубоколежащие корни, находящиеся в более влажных слоях почвы и подпочвы, могут до некоторой степени обеспечивать влагой часть корней, сосредоточенных в верхнем иссушенном слое.
Влияние температуры на корневую систему
Рост и состояние корневой системы очень сильно зависит от температуры почвы. В исследованиях реакция корней на изменение температуры выражалась в виде смен волн ускорения и торможения роста.
Вначале, при повышении температуры, наблюдается резкое ускорение роста, иногда в 15–20 раз. Далее при стабилизации температуры рост сначала резко замедляется (фаза торможения), затем восстанавливается на новом уровне в зависимости от температуры. Имеются и некоторые конкретные данные о минимальной, оптимальной и максимальной температурах для роста корней у разных видов плодовых, ягодных и орехоплодных растений.
Например, согласно исследованиям, корни яблони проявляли некоторую жизнедеятельность при температуре 1-3°С. В почве, промерзшей на глубину 68–75 см, при этом наблюдался рост корней на глубине 105–135 см.
В другом исследовании корни плодовых растений в саду начинали расти вне зависимости от глубины их залегания при температуре 5-6°С. Наиболее сильный рост корней наблюдался у яблонь и груш при температуре 10-20°С, у вишни - при 12-18°С, у абрикоса - при 12-22°С.
В третьем исследовании корни наиболее сильно росли при температуре от +7 до +20 °С, слабо - от 0 до +7°С и от +20 до +30 °С. Не наблюдался рост корней при температуре ниже 0 и выше +30 °С. Зимой слабый рост корней обнаруживался при температуре от +1,7 до +7,2°С в незамерзшем слое почвы.
Таким образом, рост корней очень тесно связан с температурой почвы. Минимальной температурой для роста корней яблони надо, по-видимому, считать +4-5°С.
Данных о максимальной температуре для роста корней очень мало. Ясно одно, максимальная температура, для разных плодовых, ягодных и орехоплодных растений находится в пределах +30-35°С.
При температурах выше +35 °С и недостатке влаги наступает быстрый процесс «суберинизации» корней (опробковение отложение в клеточной стенке жироподобного аморфного вещества суберина, в результате чего стенка клетки становится непроницаемой для жидкостей и газов).
Оптимальная температура для роста и жизнедеятельности корней, по исследовательским данным, +15–25,5 °С для яблони.
Данные о влиянии низких температур на рост и развитие корней показали причину, с чем связано наблюдающиеся нередко зимние повреждения или даже вымерзание корней. Оказалось, что очень низкой морозостойкостью при исследованиях обладали корневые системы европейских клоновых карликовых подвоев для яблони. У них уже при температуре −10...-11°С наблюдалось массовое подмерзание корней, при температуре −12°С корневая система гибла.
В последние годы было получено много новых советских и российских таких клоновых подвоев, выдерживающих без подмерзания корневой системы температуру −14...-16°С. Большей морозостойкостью корневой системы отличаются сеянцы сибирской яблони, которые переносили понижение температуры до −24,5°С.
Еще большей морозостойкостью обладает корневая система песчаной вишни. Приведенные данные морозостойкости относятся в основном к проводящим корням. По исследованию морозоустойчивости поглощающих корней получено, что наиболее устойчивыми были поглощающие корни черной смородины, малины и земляники, менее морозоустойчивыми - яблони.
Рост корней в течение вегетационного периода
Годичный рост корней включает в себя две составляющие: удлинение уже имеющихся корней и заложение новых боковых корней и последующее их удлинение. В умеренной климатической зоне растяжение корней начинается рано весной и продолжается до конца осени, дольше, чем удлинение побегов того же плодового, ягодного или орехоплодного растения.
Период между прекращением удлинения побегов и окончанием прироста корней сильно различается у разных видов указанных растений и в основном определяется климатическими особенностями сезонов года.
Многолетние исследования многих специалистов показали, что в течение вегетационного периода в зависимости от подвоя, привоя (сорта), возраста и урожайности плодового дерева и почвенно-климатических условий корни имеют от одной до нескольких, чаще всего две (весенняя и осенняя) волны роста.
Весной корни растут сильно, но после того как запасенные продукты фотосинтеза будут израсходованы для роста побегов, листьев и цветков рост корней уменьшается. При этом всасывающие корни весной больше растут в верхнем слое почвы, благодаря лучшей его прогреваемости, влажности и аэрации, а летом, наоборот, из-за сухости верхнего слоя лучше растут в более глубоком слое.
В средней, да реже и в северной зонах рост корней, например яблони, в некоторые снежные зимы, особенно после выпадения снега на незамерзшую почву, продолжается до конца декабря - и, были случаи, даже до середины января, пока температура воздуха в почве вокруг корней не опустится до 2-0°С.
В южной зоне корни плодовых, ягодных и орехоплодных растений в отдельные зимы в отдельных районах продолжают рост в течение всей зимы. Следовательно, корневая система указанных растений не имеет органического покоя, а обычно вследствие неблагоприятных условий (суровая малоснежная зима, засуха) может находиться в состоянии вынужденного покоя.
Исследования в средней зоне показали, что при оптимальных условиях водного режима корни яблони растут сравнительно равномерно в течение всего вегетационного периода, причем активные корни при этом составляют 48–70% от длины всей корневой системы дерева.
При отсутствии орошения и в обычные годы корни растут волнами. Продолжительнее и равномернее корни растут у деревьев и кустов в неурожайные или малоурожайные годы, у деревьев и кустов до начала плодоношения и в молодом возрасте с сильными приростами, а также при своевременном и хорошем уходе (удобрении, орошении и мульчировании, борьбе с болезнями и вредителями).
Так, в одном из наблюдений за деревьями яблони было обнаружено, что у деревьев с большим урожаем активные корни летом не росли в течение трех месяцев, а у деревьев без урожая - только около месяца. В данном случае вырабатываемые листьями продукты фотосинтеза на дереве с урожаем, скорее всего, расходовались исключительно на создание плодов, поэтому роста активных корней долго не было.
Корневая система плодовых, ягодных и орехоплодных растений, как показали исследования, обладает способностью в любой момент ослабленного роста перейти в состояние интенсивного роста, если для нее создадутся благоприятные условия аэрации, влажности, температуры и питания.
Многолетние исследования в разных по почвам, климату, видах растений и агротехнике зонах садоводства России показали, что весной рост корней происходит всегда, а в остальные периоды года его может не быть, что зависит от ряда условий. Может происходить сравнительно равномерный рост корневой системы от весны до зимы, а на юге при мягких зимах - весь год.
Так, в исследованиях в Московской области рост корней плодоносящих деревьев яблони сорта Антоновка на сеянцах Аниса продолжался ежегодно 6-7 месяцев, а один год, когда снег выпал на незамерзшую почву, продолжительность периода роста была 9 месяцев.
Рост корней проходил волнами: за 10 лет по две волны было 4 года, по три - 4 года, по одной - 1 год и по 4 волны - также 1 год. Максимальный рост корней в некоторые годы при орошении достигал 30–90%, а без орошения - 18–70% от суммарной длины всей корневой системы яблони. Более сильный рост активных корней 5 лет был осенью и 5 лет весной. В засушливую осень без полива рост корней может отсутствовать, а во влажную осень или при поливах быть даже более мощным, чем весной.
Зависимость от прошлогодних запасов
Рост и развитие дерева или куста весной целиком зависит от прошлогодних запасов продуктов фотосинтеза, накопленных в стволе, ветвях и корнях. С момента же появления листьев, когда исчерпываются запасенные продукты фотосинтеза, должны начать работать корни, чтобы снабжать листья в первую очередь водой и питательными веществами.
Садовод имеет большие возможности соответствующей агротехникой своевременно способствовать созданию массы активных корней и удлинению периода их роста, чтобы они росли не 4-5, а 9 месяцев. Это важно еще и потому, что активные корни осенне-зимнего периода устойчивее весенних, долговечнее и богаче водой и питательными веществами. Поэтому чем их осенью будет больше, тем легче дереву или кусту образовать большее число новых корней весной.
Отсюда становится абсолютно ясно, что чем дольше на протяжении года растут активные корни и чем их больше, тем легче дереву или кусту создавать высокие урожай. По количеству и состоянию проводящих желтых и активных белых корней можно быстро и точно диагностировать жизнедеятельность корневой системы в любое время года.
Благоприятные почвенные условия
Чтобы плодовые, ягодные и орехоплодные растения нормально росли и ежегодно давали хорошие урожаи плодов и ягод, необходимо создать им благоприятные почвенные условия для активного и длительного роста корней, то есть для обеспечения потребности данных растений во влаге и питательных веществах. Особенно важна осенняя обработка почвы, так как взрыхленная с осени почва хорошо поглощает талые воды.
Для установления возможной глубины обработки почвы в садах необходимо учитывать характер развития корневой системы у указанных растущих растений и ее размещение в почвенных горизонтах, а также свойства почвы (глубину гумусового горизонта).
Любая обработка почвы, а также агротехнические приемы, связанные с выращиванием и выкопкой саженцев и пересадкой плодовых, ягодных и орехоплодных растений, сопровождаются повреждением или потерей какой-то части их корневой системы. Однако поврежденные или утраченные части корневой системы способны восстанавливаться.
Регенерация корней
Способность к восстановлению (регенерации) - уникальное свойство корней. Регенерация может происходить на месте утраченной части корня. В садоводстве обрезка корней сопровождает почти каждый агротехнический прием, связанный с пересадкой и обработкой почвы (пикировка сеянцев, выкопка, укорачивание корней подвоев и саженцев при сортировке, обработка почвы в приствольных кругах и в междурядьях). На месте среза у одревесневших корней образуется каллус (сначала в виде кольца, а позднее он полностью закрывает рану и суберинизуется).
Под суберинизированным слоем камбиальные клетки начинают усиленно делиться и дают начало образованию новых корней по окружности. Повышение камбиальной активности связано с образованием в результате повреждения раневых гормонов (раневой кислоты), обладающих ауксиновой способностью.
Корни, имеющие гладкий срез, быстрее регенерируют на второй год. Толщина восстановленных корней независимо от диаметра среза не превышает 3-5 мм. Вместо одного корня диаметром около 20 мм появляются до 16 новых корней, но общая длина их не всегда достигает длины срезанной части. Поэтому судить о степени восстановления по числу вновь образующихся корней нельзя, необходимо измерять их длину.
Любое повреждение корневой системы, связанное с потерей части ее корней, ведет к понижению жизнедеятельности дерева или куста и снижению их последующего роста. Сильные повреждения корней особенно возможны при глубокой обработке почвы, когда перерезается много достаточно толстых одревесневших корней.
Очень негативно, как показали наблюдения, влияют даже только просто порезы корней на развитие плодовых деревьев, больных хлорозом, розеточностью, суховершиностью или имеющих ослабленный рост. В результате повреждения корней при глубокой обработке состояние указанных деревьев ухудшается еще в большей степени, усиливается поражаемость деревьев черным раком и другими болезнями и вредителями. Особенно это относится к маломощным почвам с неблагоприятными свойствами (избыточная влажность, очень высокая кислотность и другое).
Глубина обработки почвы
Рост и формирование корневой системы
Прорастающее семя образует из зародышевого корешка главный, или первичный корень, после чего вытягивается подсемядольное колено (гипокотиль), разрастаются семядоли и трогается в рост почечка (дающая стебель). В этом проявляется биполярность зародыша растения, то есть способность формировать одновременно разные органы в двух противоположных направлениях. Вскоре главный корень (нулевого порядка) удлиняется и, достигнув примерно 10-20 см, начинает ветвиться. У сеянцев плодовых растений за вегетационный период образуется до 5-7 порядков ветвления корней с преобладанием 3-4 порядка.
По данным исследований, в условиях средней зоны за вегетационный период у однолетнего сеянца яблони китайской образуется до 40 тысяч корней, имеющих суммарную длину до 230 метров. Однолетние сеянцы яблони лесной и сибирской, груши дикой, вишни и других видов плодовых развивали за это время гораздо меньше корней (по длине и количеству), по сравнению с сеянцами яблони китайской. Длина всех корней взрослого плодового дерева выражается десятками километров, а суммарное количество - миллионами отдельных корешков. Подавляющая часть корней имеет малую длину. Например, по результатам одного из исследований, у яблони корни длиной от долей миллиметра до 5 мм составляют 65,1%, у груши - 43%; корни длиной от 6 до 10 мм - соответственно 18,7 и 23%. Остальные корни имеют длину от 1-2 см до 0,5 м, но их гораздо меньше (у яблони 16,2%). Средняя длина корня у однолетних сеянцев плодовых растений довольно постоянна: у яблони - 6-7 мм, у груши - 8-9, у вишни - 8 мм. У плодоносящих деревьев плодовых растений средняя длина корня в 2 раза меньше (у яблони 3,5 мм), чем у однолетних сеянцев.
В процессе жизни растения у его корневой системы происходит циклическая смена (самоизреживание) отдельных корней. Проведенные исследования позволили установить, что у каждого растения с первых дней и до конца жизни происходит неуклонное и последовательное отмирание концов осевых корней, а также коротких боковых корней сначала на главном корне, затем по мере роста на более длинных корнях последующих порядков ветвления (первом, втором и так далее). Далее наблюдается отмирание целых мочек (сеток) корней и замена их новыми, то есть очищение (оголение) верхних, уже более толстых корней от отдельных более мелких корней и мочек.
На однолетних сеянцах за один сезон могут отмирать десятки тысяч, а у взрослых деревьев - миллионы корней. Это свидетельствует о недолговечности всасывающих корней, которые в зависимости от расположения в корневой системе живут от нескольких дней до месяца и редко дольше. Отмирание корней было названо одним из ведущих исследователей корневой системы плодовых и ягодных растений в период СССР В. А. Колесниковым корнепадом. Это название достаточно прижилось и используется и сейчас. Оно несколько созвучно с явлением листопада у плодовых, ягодных и орехоплодных растений.
У разных видов растений отмирание корней может ускоряться или замедляться в разной степени, в зависимости от условий жизни. В результате отмирания и возобновления корневая система растения непрерывно в течение вегетации и всей жизни продвигается во все новые, неиспользованные слои почвы. Она распространяется от ствола круговыми полосами, отмирая в одном месте и появляясь в другом, и обеспечивает нормальные условия роста и плодоношения растений. Суммарная масса отмерших в течение года корней выражается десятками килограммов и даже тоннами органических остатков в расчете на гектар, являющихся источниками питания микроорганизмов ризосферы. В процессе корнепада происходит отмирание не только обрастающих (мелких корней), но и скелетных. Отмирание и возобновление корней, или циклическая смена их, у сеянцев и взрослых плодовых, ягодных и орехоплодных растений представляет естественный процесс в жизненном цикле их развития и свойственно всем древесным и травянистым растениям.
Рост и формирование корневой системы в почве зависит от происхождения корней (семенные или вегетативные), вида и сорта растения (привой), природных условий и агротехники. Корневые системы семенного происхождения обычно проникают в почву глубже, чем вегетативного происхождения. Различны типы роста корней в стороны и в глубь почвы, а также их расположение по горизонтали почвы и почвогрунта. Например, в средней полосе ежегодный прирост корней плодовых растений в сторону от ствола по радиусу составлял около 20-36 см, а в глубину - 17-33 см. Различия значительны, и садовод должен учитывать их при выборе и подготовке почвы под сад и при дальнейшем уходе за корнями и почвой. Агротехническими приемами, например плантажной обработкой и удобрением, на любой почве, в том числе и на распространенной у нас дерново-подзолистой, можно увеличить массу корней, расположенных ниже пахотного горизонта. Тонкие, обрастающие и всасывающие корни в зависимости от природных условий и агротехники могут расти с ранней весны и до поздней осени, даже после листопада, а затем прекращать рост и отмирать. В общем, постепенно разрастаясь, корневая система сеянца или саженца, а позднее взрослого плодового, ягодного или орехоплодного растения охватывает огромные объемы почвы и почвогрунта длинными и толстыми тяжами корней, на которых размещены сотни тысяч коротких корешков, а на них десятки миллионов коротких волосков.
Корни горизонтального направления у плодовых пород в северной зоне в массе располагаются на глубине примерно до 30-50 см, в средней - до 50-75 см, у большинства ягодных пород в этих же зонах - на глубине 0-60 см, у карликовых деревьев яблони - на глубине 0-50 см, у стланцевых деревьев яблони и груши - на глубине 0-40 см. В результате наличия плотных, а местами сцементированных прослоек и погребенных горизонтов в почве корневая система может залегать ярусами. Например, в Среднем Поволжье на дерново-подзолистой почве первый ярус корней яблони залегает на глубине 18 см, второй - 101-140 см. Более тонкие корни в значительном количестве могут встречаться во всех зонах садоводства в поверхностном слое почвы с глубины 3-10 см. У яблони в северной зоне корни чаще размещены глубже 10-20 см от поверхности почвы.
Проведенные исследования показали, что у плодовых растений, начиная со второго года и всю последующую жизнь, диаметр корневой системы в 1,5-2 раза больше диаметра (проекции) кроны. Такое соотношение диаметра крон и корневых систем наблюдается у подавляющего большинства плодовых, ягодных и орехоплодных растений во всех зонах садоводства. Так, опыты, проведенные в средней полосе, показали, что у 7-летних деревьев яблони корневая система разрастается в стороны по диаметру на 3,5 м, 14-летних - на 5 м, у 20-летних - на 8-9 м. При этом выявился следующий порядок распределения разных видов плодовых растений по диаметру распространения корневой системы - абрикос, яблоня, груша, слива, вишня.
Корни вертикального направления у разных плодовых, ягодных и орехоплодных растений проникают в северной зоне в большинстве случаев до 1-2 м, в средней - до 2-4 м, а в южной - до 5-10 м. Вглубь корни продвигаются главным образом по ходам землероев, особенно дождевых червей, и по трещинам почвы. На глубину проникновения корней сильное влияние оказывают почвенные условия, привой и подвой.
Так, на дерново-подзолистой почве в Московской области корни яблони проникают на глубину до 3-4 м, груши, сливы и вишни - до 2 м. На этой же почве основная масса вертикальных корней Антоновки обыкновенной на китайке залегает на глубине 1 м, на полукарликовом клоновом подвое - 1,3 м, на яблоне лесной и сеянцах Аниса - 2 м, а на сеянцах Антоновки - на глубине 2,5 м. Количество и длина корней вертикального направления может быть значительным, особенно у плодовых деревьев, выращиваемых в южной зоне. В этой зоне корни даже карликовых клоновых подвоев могут достигать глубины 5 м.
Как правило, чем южнее произрастают садовые растения, тем глубже залегают их корневые системы. Однако и на юге корневые системы могут быть расположены поверхностно. Это наблюдается в случаях близкого залегания грунтовых вод или плотных, а иногда галечных прослоек и сцементированных отложений. В таких случаях данные растения оказываются менее урожайными и менее долговечными. Например, в Крыму деревья яблони сорта Сары Синап в возрасте 35 лет при размещении корней на глубине до 4,6 м давали до 300 кг плодов, а деревья того же сорта и возраста при других почвенных условиях (близкие грунтовые воды) и глубине залегания корней до 1,5 м давали только 100 кг.
Глубина проникновения корней в почву имеет очень большое значение. Специалисты указывают, что деревья яблони в средней зоне, имеющие глубину залегания корней до 3 м и больше, гораздо лучше развивались и плодоносили, чем деревья с глубиной проникновения корней до 1,6 м, и что деревья яблони с поверхностными корнями (мочкой) больше нуждаются в удобрении и болезненно реагируют на всякие недочеты в агротехнике во время вегетации. Кроме того, крупные корни при поверхностном их залегании будут ежегодно повреждаться при обработке почвы, тонкие же корни намного хуже будут обеспечены в верхнем слое почвы влагой, что может привести к их отмиранию при отсутствии регулярных поливов. С более глубоким залеганием корневой системы связан и меньший выпад плодовых деревьев. Так, по наблюдениям в яблоневом саду в средней зоне при паровой обработке почвы, когда у деревьев была глубокая корневая система, общий выпад деревьев за 13 лет после посадки составлял 2,4% против 11,1% у деревьев с менее глубоко залегающей корневой системой при задернении почвы в саду.
Лучшей корневой системой садовых растений следует считать такую, которая залегает равномерно по кругу, более глубоко и широко и имеет возможность извлекать воду и питательные вещества из большого объема почвы и почвогрунта. Все указанные растения в этом случае характеризуются большей засухо- и морозоустойчивостью.
Исследованиями отечественными и зарубежными учеными установлено, что данные растения, особенно плодовые, с мощной корневой системой гораздо дольше живут, более зимостойки, лучше и регулярнее плодоносят. Поэтому очень важно соответствующими агротехническими приемами способствовать глубокому залеганию корневой системы. Вместе с тем садовод всегда перед началом обработки почвы должен проверять глубину залегания корней, чтобы правильно назначить на каждом садовом участке под каждым деревом и кустом глубину обработки, внесения удобрений и тому подобное.
При выращивании в саду плодовых, ягодных и орехоплодных растений наблюдается взаимовлияние их корневых систем. Особенно такое влияние (конкуренция) корней наблюдалась между разными экземплярами растений одного и того же вида. При этом корни растений распространяются в направлении слабого соседнего растения. При наблюдении за корневыми системами разных плодовых растений было отмечено:
1. Корни яблони в одних местах не входят, а в других входят в корни соседней яблони.
2. Корни яблони входят в корневую систему выкорчеванного в прошлом году дерева яблони, но уходят от корневых систем сеянцев яблони только что выкорчеванного питомника.
3. Корни сливы, черешни свободно входят в корневую систему яблони, То же самое наблюдалось в отношении корней яблони, груши и абрикоса, миндаля, то есть между корнями этих пород нет конкуренции ни за пищу, ни за воду и они не испытывают никакого неудобства из-за присутствия корней растений других видов.
Проведенные исследования показали, что главную роль в совместимости корневых систем растений одного вида или разных видов играет аллелопатическое их действие посредством выделения в почву токсических веществ (колинов). Проявление корневыми системами плодовых, ягодных и орехоплодных растений явления аллелопатии следует обязательно учитывать при их выращивании.
Взаимосвязь роста корневой системы от внешних условий
Проведенными наблюдениями между ростом корневой и надземной систем плодовых, ягодных и орехоплодных растений обнаружена определенная взаимосвязь. Оказалось, например, что присутствие растущих корешков на корнях саженцев этих растений значительно сказывается на приросте их побегов. Так, при наличии растущих корешков рост побегов после посадки саженцев яблони в сад был более сильным (413 см на одно растение против 148 см). Изучение суточного прироста корней и плодов у яблони сорта Славянка показало наличие и между ними определенной взаимосвязи при усилении роста корней ослабевал рост плодов, а затем происходило обратное. Проводились и наблюдения за установлением соотношения в процессе роста плодовых, ягодных и орехоплодных растений между размерами корней и ветвей. Такие наблюдения показали, что, например, у молодых растений яблони, груши, сливы и других прирост корней неизменно превышал прирост ветвей, причем в весьма значительной степени. А в последующие годы, наоборот, более усиленно происходил рост надземных органов.
Общепринятым обычно раньше считали наличие автономной связи между отдельными крупными скелетными корнями и отдельными скелетными ветвями. Однако, начиная с конца 50-х годов прошлого века, благодаря использованию для изучения процессов питания растений метода меченых атомов, были получены данные, которые не полностью согласуются с этим общепринятыми когда-то взглядами. Так, в одном из опытов автономная связь между корнями и ветвями наблюдалась только у корнесобственных яблонь, но у привитых яблонь такой автономной связи не наблюдалось. В другом опыте указывалось, что наряду с локализацией основного количества радиофосфора в одной из скелетных ветвей кроны наблюдалось распространение небольших его количеств и в остальной части кроны. Поэтому в настоящее время считают, что строгой автономной связи между отдельными основными крупными скелетными корнями и основными скелетными ветвями не существует. Говорят лишь о наличии относительной автономной связи.
Для роста корней очень большое значение имеет здоровый листовой аппарат. Так, в исследованиях летнее удаление боковых побегов приводило к задержке роста корней яблони. Корневая система начинает хуже развиваться при нарушении нормальной деятельности листового аппарата, что бывает, например, при неправильной обрезке растений, повреждении листьев вредителями и болезнями, преждевременном листопаде и так далее. Рост корней у плодовых деревьев с урожаем бывает более слабый, чем у деревьев без урожая. Отличается и сам характер роста корней - у деревьев с урожаем наблюдается медленный рост, быстрое побурение многих боковых разветвлений, более коротких, чем у деревьев без урожая. Отставание роста корней на деревьях с урожаем обусловлено большим расходом питательных веществ на рост плодов, то есть тем же, чем объясняется слабый прирост побегов в урожайные годы. Улучшить рост активных корней в урожайные годы и вместе с тем ослабить склонность растений к периодичности плодоношения можно хорошо известными способами - внесением удобрений, поливом, прореживанием цветков и надлежащей обрезкой.
Были проведены и исследования по развитию корневой системы яблони в зависимости от развития надземной части. Оказалось, что существует определенная взаимосвязь между развитием всасывающей поверхности корня сеянца яблони и развитием листового аппарата. Отношение всасывающей поверхности корня к площадям листового аппарата оставалось почти постоянным в течение всей вегетации до периода листопада и равнялось 1,5-2.
На рост корневой системы очень сильное влияние оказывают внешние условия, а также приемы агротехники, в первую очередь связанные с воздействием на почву (обработка почвы, внесение удобрений, полив, борьба с болезнями и вредителями и так далее). Особенно большое влияние оказывает на развитие корневой системы аэрация почвы, водный, температурный и пищевой режимы почвы.
Значение аэрации для роста корней было показано во многих исследованиях. Эти исследования показали, что если около дерева или куста почва была разной плотности, то корни развивались в сторону рыхлой почвы. На более плотных почвах наблюдалось и более поверхностное развитие корневой системы. При лучшей аэрации наблюдалась и большая разветвленность корней. В одном из исследований в условиях вегетационного опыта при недостатке кислорода резко ослаблялось развитие и корневой системы, и надземных органов (появление признаков хлороза, усыхание листьев и другое). Особенно резко реагировали на аэрацию корневые волоски. Минимальным содержанием кислорода в данном опыте для корневых волосков яблони является 7,5 мл в литре воды, а для смородины - 5,8 мл. Важное значение имеет аэрация и для нормального питания. При недостатке в почвенном воздухе кислорода происходило нарушение поступления минеральных веществ в сеянцы яблони.
В другом вегетационном опыте однолетки яблони были посажены в сосуды, набитые почвой разной степени уплотнения, а также почвой, смешанной с песком. При этом оказалось, что количество новообразовавшихся корней было значительно большим в рыхлой почве, а также в почве, смешанной с песком, и значительно меньшим в уплотненной почве.
По данным исследований корни проникают свободно в почву с плотностью ниже 30 кг на 1 кв. см, гораздо хуже при плотности 30-60 кг на 1 кв. см и совсем не растут при плотности выше 60 кг на 1 кв. см. Совершенно непроницаемыми для корней плодовых деревьев являются сильно сцементированные прослойки с плотностью 300 кг на 1 кв. см.
Интерес представляет и влияние на корневую систему составных частей почвенного воздуха - кислорода, углекислоты и азота. В исследованиях дыхание корней яблони ослаблялось при уменьшении содержания кислорода в почвенном воздухе ниже 8-9%, а также при повышении содержания угольной кислоты выше 5-6%. Недостаток кислорода особенно сказывался на состоянии корневых волосков: достаточно было 30-минутного пребывания всасывающего корня в бескислородной среде, чтобы корневые волоски не образовывались. Плодовые, ягодные и орехоплодные растения особенно чувствительны к недостатку кислорода в периоды сильного роста активных корней (весной и осенью). Больше кислорода требовалось для развития кончиков корней (5-10%) и еще больше для появления новых корней (12%). При содержании кислорода менее 10% снижался сухой вес корней, а ниже 15% - ослаблялось поглощение и накопление зольных элементов в растении. У большинства растений замена в почвенном воздухе кислорода азотом приводит к уменьшению всасывающей способности корней, а в дальнейшем - к их отмиранию.
Таким образом, для корней плодовых, ягодных и орехоплодных растений в почвенном воздухе обязательно необходимо присутствие кислорода, в то же время избыток углекислоты вредит им больше, чем недостаток кислорода. При нормальном газообмене между почвой и атмосферой почвенный воздух наполняется кислородом и освобождается от избытка угольной кислоты, то есть процесс идет именно так, как это необходимо для лучшего развития корней.
Большую роль в росте корневой системы играет влажность почвы. Достаточная влажность почвы является особенно важной для активных мочковатых корней, которые развиваются лишь в достаточно увлажненных слоях почвы. На сухих мало орошаемых почвах корни развиваются больше в горизонтальном направлении, чем на почвах достаточно увлажненных. Такое же поверхностное залегание корней наблюдается и при дерновой системе содержания почвы, например в яблоневом саду, по сравнению с паром. Очень большое значение для роста корневой системы имеет полив.
Так, в одном из опытов без орошения длина скелетных корней шестилетнего дерева яблони составляла 15-58 м, то при орошении она равнялась 3219 м. Под влиянием орошения увеличилась также глубина залегания и распространения корней в горизонтальном направлении. В другом опыте с корневой системой яблони было получено, что она намного лучше развивалась, когда поливы делались при более высокой влажности, то есть когда поливами поддерживалась в течение всего вегетационного периода большая влажность почвы.
Отрицательное влияние оказывает на рост корней и избыток влаги, в связи с ухудшением условий аэрации. Этим, кстати, объясняется угнетающее влияние на корневую систему неглубокого залегания грунтовых вод.
К сожалению, я не нашел точных наблюдений о влиянии на рост корней разной степени увлажнения почвы. Но в целом, опытные данные показывают, что ослабление роста корней начинается задолго до начала завядания растений. Так, в одном из исследований рост корней и надземных частей начал замедляться при постепенном нарастании засушливых условий за 7-11 суток до начала увядания листьев. Поглощающие корни чувствительнее к недостатку влаги, чем проводящие - при длительной засухе они прекращали рост за 1-3 суток до начала увядания листьев. Прекращение роста этих корней сопровождается быстрой их субернизацией, и вслед за этим или одновременно начинается увядание листьев. Отрицательно реагируют поглощающие корни и на воздушную засуху. В этом же исследовании рост их прекращался даже при кратковременном снижении влажности воздуха с 90-80 до 60-50%. Однако, хотя рост активных корней при недостатке влаги и прекращается, они могут до 3-4 месяцев оставаться живыми, не теряя способности частичного всасывания, и при благоприятных условиях восстанавливают свою активность.
Интересные данные были получены в исследовании о возможности передвижении влаги в самом растении в те участки корневой системы, которые испытывают недостаток в ней. Обнаружилось, что корни, находящиеся в сухой почве, могут частично получать влагу от корней, находящихся в более увлажненных слоях почвы. Последним можно объяснить эффективность чересполосного задернения междурядий, хороший рост и высокую урожайность плодовых деревьев при глубоком залегании корней. Глубоколежащие корни, находящиеся в более влажных слоях почвы и подпочвы, могут до некоторой степени обеспечивать влагой часть корней, сосредоточенных в верхнем иссушенном слое.
Рост и состояние корневой системы очень сильно зависит от температуры почвы. В исследованиях реакция корней на изменение температуры выражалась в виде смен волн ускорения и торможения роста. Вначале при повышении температуры наблюдается резкое ускорение роста, иногда в 15-20 раз. Далее при стабилизации температуры рост сначала резко замедляется (фаза торможения), затем восстанавливается на новом уровне в зависимости от температуры. Имеются и некоторые конкретные данные о минимальной, оптимальной и максимальной температурах для роста корней у разных видов плодовых, ягодных и орехоплодных растений.
Так, по одному из исследований, корни яблони проявляли некоторую жизнедеятельность при температуре 1-3°C . В почве, промерзшей на глубину 68-75 см, при этом наблюдался рост корней на глубине 105-135 см. Во втором исследовании корни плодовых растений в саду начинали расти вне зависимости от глубины их залегания при температуре 5-6°C . Наиболее сильный рост корней наблюдался у яблонь и груш при температуре 10-20°C , у вишни - при 12-18°C , у абрикоса - при 12-22°C . В третьем исследовании корни наиболее сильно росли при температуре от 7 до 20°C , слабо - от 0 до 7°C и от 20 до 30°C . Не наблюдался рост корней при температуре ниже 0 и выше 30°C . Зимой слабый рост корней обнаруживался при температуре от 1,7 до 7,2°C в незамерзшем слое почвы.
Таким образом, рост корней очень тесно связан с температурой почвы. Минимальной температурой для роста корней яблони надо, по-видимому, считать 4-5°С. Данных о максимальной температуре для роста корней имеется мало. Очевидно, она лежит для разных плодовых, ягодных и орехоплодных растений где-то в пределах 30-35°C . При более высоких температурах, а также недостатке влаги наступает быстрый процесс «суберизации» корней. Оптимальная температура для роста и жизнедеятельности корней, по данным ряда исследователей, 15-25,5°C для яблони.
Представляют также интерес данные о влиянии на рост корней низких температур, с чем связано наблюдающиеся нередко зимние повреждения или даже вымерзание корней. Оказалось, что очень низкой морозостойкостью при исследованиях обладали корневые системы европейских клоновых карликовых подвоев для яблони. У них уже при температуре -10...-11°C наблюдалось массовое подмерзание корней, при температуре -12°C корневая система гибла.
В последние годы было получено много новых советских и российских таких клоновых подвоев, выдерживающих без подмерзания корневой системы температуру -14...-16°C
. Большей морозостойкостью корневой системы отличаются сеянцы сибирской яблони, которые переносили понижение температуры до -24,5°C
. Еще большей морозостойкостью обладает корневая система песчаной вишни. Приведенные данные морозостойкости относятся в основном к проводящим корням. По исследованию морозоустойчивости поглощающих корней получено, что наиболее устойчивыми были поглощающие корни черной смородины, малины и земляники, менее морозоустойчивыми - яблони.
Сезонная динамика роста корней
Годичный рост корней включает в себя две составляющие: удлинение уже имеющихся корней и заложение новых боковых корней и последующее их удлинение. В умеренной климатической зоне растяжение корней начинается рано весной и продолжается до конца осени, дольше, чем удлинение побегов того же плодового, ягодного или орехоплодного растения. Период между прекращением удлинения побегов и окончанием прироста корней сильно различается у разных видов указанных растений и в основном определяется климатическими особенностями сезонов года.
Многолетние исследования многих специалистов показали, что в течение вегетационного периода в зависимости от подвоя, привоя (сорта), возраста и урожайности плодового дерева и почвенно-климатических условий корни имеют от одной до нескольких, чаще всего две (весенняя и осенняя) волны роста. Весной корни растут сильно, но после того как запасенные продукты фотосинтеза будут израсходованы для роста побегов, листьев и цветков рост корней уменьшается. При этом всасывающие корни весной больше растут в верхнем слое почвы, благодаря лучшей его прогреваемости, влажности и аэрации, а летом, наоборот, из-за сухости верхнего слоя лучше растут в более глубоком слое. В средней, да реже и в северной зонах рост корней, например яблони, в некоторые снежные зимы, особенно после выпадения снега на незамерзшую почву, продолжается до конца декабря - и, были случаи, даже до середины января, пока температура воздуха в почве вокруг корней не опустится до 2-0°С. В южной зоне корни плодовых, ягодных и орехоплодных растений в отдельные зимы в отдельных районах продолжают рост в течение всей зимы. Следовательно, корневая система указанных растений не имеет органического покоя, а обычно вследствие неблагоприятных условий (суровая малоснежная зима, засуха) может находиться в состоянии вынужденного покоя.
Исследования в средней зоне показали, что при оптимальных условиях водного режима корни яблони растут сравнительно равномерно в течение всего вегетационного периода, причем активные корни при этом составляют 48-70% от длины всей корневой системы дерева. При отсутствии орошения и в обычные годы корни растут волнами. Продолжительнее и равномернее корни растут у деревьев и кустов в неурожайные или малоурожайные годы, у деревьев и кустов до начала плодоношения и в молодом возрасте с сильными приростами, а также при своевременном и хорошем уходе (удобрении, орошении и мульчировании, борьбе с болезнями и вредителями). Так, в одном из наблюдений за деревьями яблони было обнаружено, что у деревьев с большим урожаем активные корни летом не росли в течение трех месяцев, а у деревьев без урожая - только около месяца. В данном случае вырабатываемые листьями продукты фотосинтеза на дереве с урожаем, скорее всего, расходовались исключительно на создание плодов, поэтому роста активных корней долго не было.
Корневая система плодовых, ягодных и орехоплодных растений, как показали исследования, обладает способностью в любой момент ослабленного роста перейти в состояние интенсивного роста, если для нее создадутся благоприятные условия аэрации, влажности, температуры и питания. Многолетние исследования в разных по почвам, климату, видах растений и агротехнике зонах садоводства России показали, что весной рост корней происходит всегда, а в остальные периоды года его может не быть, что зависит от ряда условий. Может происходить сравнительно равномерный рост корневой системы от весны до зимы, а на юге при мягких зимах - весь год.
Так, в исследованиях в Московской области рост корней плодоносящих деревьев яблони сорта Антоновка на сеянцах Аниса продолжался ежегодно 6-7 месяцев, а один год, когда снег выпал на незамерзшую почву, продолжительность периода роста была 9 месяцев. Рост корней проходил волнами: за 10 лет по две волны было 4 года, по три - 4 года, по одной - 1 год и по 4 волны - также 1 год. Максимальный рост корней в некоторые годы при орошении достигал 30-90%, а без орошения - 18-70% от суммарной длины всей корневой системы яблони. Более сильный рост активных корней 5 лет был осенью и 5 лет весной. В засушливую осень без полива рост корней может отсутствовать, а во влажную осень или при поливах быть даже более мощным, чем весной.
Рост и развитие дерева или куста весной целиком зависит от прошлогодних запасов продуктов фотосинтеза, накопленных в стволе, ветвях и корнях. С момента же появления листьев, когда исчерпываются запасенные продукты фотосинтеза, должны начать работать корни, чтобы снабжать листья в первую очередь водой и питательными веществами. Садовод имеет большие возможности соответствующей агротехникой своевременно способствовать созданию массы активных корней и удлинению периода их роста, чтобы они росли не 4-5, а 9 месяцев. Это важно еще и потому, что активные корни осенне-зимнего периода устойчивее весенних, долговечнее и богаче водой и питательными веществами. Поэтому чем их осенью будет больше, тем легче дереву или кусту образовать большее число новых корней весной. Отсюда становится абсолютно ясно, что чем дольше на протяжении года растут активные корни и чем их больше, тем легче дереву или кусту создавать высокие урожай. По количеству и состоянию проводящих желтых и активных белых корней можно быстро и точно диагностировать жизнедеятельность корневой системы в любое время года.
Чтобы плодовые, ягодные и орехоплодные растения нормально росли и ежегодно давали хорошие урожаи плодов и ягод, необходимо создать им благоприятные почвенные условия для активного и длительного роста корней, то есть для обеспечения потребности данных растений во влаге и питательных веществах. Особенно важна осенняя обработка почвы, так как взрыхленная с осени почва хорошо поглощает талые воды. Для установления возможной глубины обработки почвы в садах необходимо учитывать характер развития корневой системы у указанных растущих растений и ее размещение в почвенных горизонтах, а также свойства почвы (глубину гумусового горизонта). Любая обработка почвы, а также агротехнические приемы, связанные с выращиванием и выкопкой саженцев и пересадкой плодовых, ягодных и орехоплодных растений, сопровождаются повреждением или потерей какой-то части их корневой системы. Однако поврежденные или утраченные части корневой системы способны восстанавливаться.
Способность к восстановлению (регенерации) - уникальное свойство корней. Регенерация может происходить на месте утраченной части корня. В садоводстве обрезка корней сопровождает почти каждый агротехнический прием, связанный с пересадкой и обработкой почвы (пикировка сеянцев, выкопка, укорачивание корней подвоев и саженцев при сортировке, обработка почвы в приствольных кругах и в междурядьях). На месте среза у одревесневших корней образуется каллус (сначала в виде кольца, а позднее он полностью закрывает рану и суберинизуется). Под суберинизированным слоем камбиальные клетки начинают усиленно делиться и дают начало образованию новых корней по окружности. Повышение камбиальной активности связано с образованием в результате повреждения раневых гормонов (раневой кислоты), обладающих ауксиновой способностью. Корни, имеющие гладкий срез, быстрее регенерируют на второй год. Толщина восстановленных корней независимо от диаметра среза не превышает 3-5 мм. Вместо одного корня диаметром около 20 мм появляются до 16 новых корней, но общая длина их не всегда достигает длины срезанной части. Поэтому судить о степени восстановления по числу вновь образующихся корней нельзя, необходимо измерять их длину.
Любое повреждение корневой системы, связанное с потерей части ее корней, ведет к понижению жизнедеятельности дерева или куста и снижению их последующего роста. Сильные повреждения корней особенно возможны при глубокой обработке почвы, когда перерезается много достаточно толстых одревесневших корней. Очень негативно, как показали наблюдения, влияют даже только просто порезы корней на развитие плодовых деревьев, больных хлорозом, розеточностью, суховершиностью или имеющих ослабленный рост. В результате повреждения корней при глубокой обработке состояние указанных деревьев ухудшается еще в большей степени, усиливается поражаемость деревьев черным раком и другими болезнями и вредителями. Особенно это относится к маломощным почвам с неблагоприятными свойствами (избыточная влажность, очень высокая кислотность и другое).
На всех почвах надо быть весьма осторожным при установлении глубины обработки в садах и на садовых участках, в которых почва в течение ряда лет была задернена, что стимулирует более поверхностное залегание корней. А сейчас на многих садовых участках в наших коллективных садах искусственное задернение почвы стало модным. Очень осторожно надо обрабатывать почву в карликовых и стланцевых садах, а также на ягодниках, где много корней залегает в поверхностных слоях. Надо принять за правило, что при обработке почвы нельзя допускать повреждения корней диаметром больше 8-10 мм (толще карандаша). Поскольку корни такого диаметра и выше на дерново-подзолистых малоструктурных почвах, которые преобладают у нас, отрастают очень слабо и во многих случаях остаются обрубками, а их повреждение отрицательно сказывается на состоянии растения. Хотя регенерация корней зависит от местных почвенных и климатических условий, а также от характера подвоев, то диаметр корней, которые нежелательно повреждать, можно быть и чуть иным (больше или меньше).
Таким образом, при решении вопроса о глубине обработки почвы надо иметь данные о глубине залегания корней толщиной 8-10 мм или другого диаметра, которые при данных условиях не следует повреждать. Поскольку глубина эта не постоянная не только для разных садов и разных садовых участков, но даже для отдельных их частей, то надо хотя бы один раз в 3-5 лет в каждом таком саду и каждом садовом участке сделать контрольные раскопки корней. Контрольную раскопку делают на площадках 1 кв. м по периферии кроны. Еще более наглядную картину можно получить, если вскрыть узкую полосу почвы шириной 30-40 см от ствола до середины междурядий на глубину залегания корней толщиной 5-10 мм. При этом обработку надо проводить на 1-2 см мельче того слоя, где залегают корни такого диаметра.
Учитывая неодинаковую глубину залегания корней на разном расстоянии от штамба, следует обрабатывать почву на меньшую глубину в приштамбовой зоне и на большую в междурядьях. Если на одной площади сада или всегда садового участка посажены разные виды плодовых, ягодных и орехоплодных растений (или привитые на разные подвои) с неодинаковой глубиной залегания корней, то глубину обработки почвы устанавливают по тому виду, сорту и подвою, у которых корни залегают наименее глубоко. В отдельные годы глубину обработки нужно несколько изменять. Поскольку если в течение ряда лет обрабатывать почву на одинаковую глубину, то образуется уплотненный слой (подошва), от чего ухудшаются условия роста корней.
Способность к регенерации и увеличению активной части корней у разных подвоев выражена неодинаково. Подвои различают по пороговому пределу толщины корней, при подрезке которых отношение длины вновь образованных корней к длине срезанных через год больше единицы. В таком случае наблюдается положительный регенерационный эффект. Когда длина восстановившихся корней не достигает длины срезанных, наблюдается отрицательный эффект, свидетельствующий о вредности обрезки корней.
В принципе, проведя на своем садовом участке подобную ориентировочную опытную обрезку разного диаметра корней у разных подвоев, можно определить и пороговый предел толщины корней, обрезка которых дает положительный эффект. А определив такой пороговый предел, можно сделать и обрезку ряда таких пороговых корней у подвоя конкретного дерева. В литературе отмечается, что положительный эффект получается при удалении до 30% суммарной длины корней у растений, привитых на семенные подвои, а у растений, привитых на вегетативные подвои, до 40%.
Лучшая регенерация корней наблюдается только при достаточной аэрации и влажности почвы, а также соответствующей температуре и наличии достаточного количества питательных веществ. Учитывая большую роль влажности в регенерации корней, основную обработку почвы следует делать при достаточной ее влажности (после полива или дождей). Очень важным фактором для регенерации корней является температура. Причем если оптимальная температура для обычного роста корней, например, яблони равна 15-20,5°C , то для лучшего образования каллуса на обрезанных корнях у этого же вида растения температура почвы должна быть 29,4-32,2°C , то есть значительно более высокой.
Очень важным для регенерации корней являются сроки обработки почвы. В опытах по изучению восстановления корней у яблони в зависимости от времени нанесения поранения, что важно для установления лучшего срока осенней обработки почвы, было установлено следующее. Восстановление корней начинается с образования каллуса через 20-30 дней после нанесения поранения.
Через 30-50 дней появляются новые корни. На корнях диаметром до 5 мм новообразование корней происходит более интенсивно и быстрее, чем на более толстых корнях. Через 50-70 дней пораненный участок корня покрывается каллусом и образуется мочка разветвленных корней, полностью покрывающих срез.
Особый интерес в этих опытах представляют наблюдения за восстановлением корней яблони в период пожелтения листьев и после листопада. На корнях, пораненных в период пожелтения листьев, к концу сентября того же года образовалось кольцо каллуса и отросли белые корни длиной 2-3 мм. Весной следующего года до набухания почек начался рост молодых корней около среза. В период сильного роста побегов в начале июня на корнях образовалось много разветвленных деятельных белых корней длиной до 1,5 м. Корни, пораненные после листопада, не покрылись каллусом до наступления устойчивых морозов, и весной следующего года на срезах наблюдалось отмирание тканей коры и загнивание их. Рост новых корней начался с запозданием лишь 15 мая, причем молодые корни появились на 1-2 см выше среза. К периоду сильного роста побегов рост корней несколько усилился, но все же отставал от роста корней, пораненных ранней осенью перед пожелтением листьев.
То же самое подтвердилось и во многих других подобных опытах, проводимых с самыми разными плодовыми, ягодными и орехоплодными растениями. Таким образом, наилучший срок осенней обработки почвы - период перед пожелтением и опадением листьев. Поскольку лучшая регенерация корней при ранних сроках обработки почвы может быть объяснена с одной стороны лучшими условиями внешней среды (влажность почвы, аэрация, температура), а с другой - наличием целых листьев, обеспечивающих образование продуктов фотосинтеза, необходимых для отрастания корней. Поэтому осеннюю обработку почвы в садах и на садовых участках можно проводить у нас в первой половине сентября после съема урожая, при полном сохранении листьев, до начала второго (осеннего) роста корней, чтобы еще до осени у деревьев и кустов успели восстановиться поврежденные корни и накопился дополнительный, необходимый для восстановления запас продуктов фотосинтеза.
Однако надо отметить, что ранняя осенняя обработка почвы в садах и на садовых участках (до листопада) имеет тот серьезный недостаток, что при ней не обеспечивается заделка в почву опавших листьев, что необходимо для борьбы с рядом опасных грибных болезней, в частности с паршой, коккомикозом и другими. Правда, возможен сбор опавших листьев и помещение их в компост или сжигание, но по обработанной почве это провести значительно труднее. Кроме того, проведение осенней обработки в ранние сроки, особенно с зимними сортами (до уборки урожая), затрудняется наличием в саду большого числа подпор и какую-то часть осенней обработки почвы приходится делать позднее.
Ни в коем случае нельзя делать осеннюю обработку почвы, когда почва иссушена, не рыхлится, иначе поверхность почвы будет глыбистой. Недопустимость такой обработки обусловлена тем, что при недостатке влаги поврежденные корни будут плохо регенерировать. Кроме того, глыбистая почва вследствие диффузного испарения будет еще в большей степени высыхать, а зимой она будет на большую глубину промерзать, что усилит опасность зимнего повреждения корней от низких температур, особенно в условиях малоснежной зимы. Поэтому, если ко времени оптимального срока осенней обработки почва иссушена, следует вместо обработки сделать мелкое рыхление, обработку перенести на несколько более поздний срок. В крайнем случае, можно даже отказаться от осенней обработки почвы, что будет лучше, чем глыбистая зябь, очень опасная для плодовых, ягодных и орехоплодных растений.
В. Н. Шаламов
Роль корней. Корни плодовых и ягодных растений выполняют следующие основные функции.
1. Всасывание из почвы воды и растворенных в ней минеральных солей и азота в виде нитратов, которые в активных корнях восстанавливаются до аммиака, используемого при образовании аминов, амидов, аминокислот и других органических соединений. Следовательно, корни, как и листья, обладают способностью вырабатывать органические вещества.
2. Активное воздействие на твердую фазу почвы и перевод в раствор части адсорбированных ионов, а также усвоение углекислоты почвы и снабжение продуктами ее ассимиляции надземных органов. При этом жизнедеятельность листьев обеспечивается водой и веществами, ассимилируемыми корнями, которые, в свою очередь, снабжаются энергией и углеводами, вырабатываемыми в листьях. Корневой системе растения свойственна функция регулирования окислительно- восстановительного режима листьев (Иванов, 1953).
3. Выделение в почвенную среду органических кислот, фосфора, калия и других веществ, способствующих растворению минеральных веществ почвы и питанию микроорганизмов ризосферы.
4. Взаимодействие (симбиоз) с микроорганизмами, а также с ризосферным комплексом микроорганизмов.
5. Накопление запасных питательных веществ.
6. Закрепление растения в устойчивом положении.
Рост корней. Прорастающее семя образует из зародышевого корешка главный, или первичный, корень, после чего вытягивается подсемядольное колено (гипокотиль), разрастаются семядоли и трогается в рост почечка (дающая стебель). В этом проявляется биполярность зародыша растения, то есть способность формировать одновременно разные органы в двух противоположных направлениях.
Вскоре главный корень (нулевого порядка) удлиняется и, достигнув примерно 10 - 20 см, начинает ветвиться. У сеянцев плодовых пород за вегетационный период образуется до 5 - 7 порядков ветвления корней с преобладанием 3 - 4-го порядка. По нашим данным, в условиях Московской области за вегетационный период у однолетнего сеянца яблони китайской образуется до 40 тыс. корней, имеющих суммарную длину до 230 м. Однолетние сеянцы яблони лесной и сибирской, груши дикой, вишни и других пород развивают за это время гораздо меньше корней (по длине и количеству) по сравнению с сеянцами яблони китайской. Длина всех корней взрослого дерева плодовых пород выражается десятками километров, а суммарное количество - миллионами отдельных корешков.
Подавляющая часть корней имеет малую длину. Например, у яблони корни длиной от долей миллиметра до 5 мм составляют 65,1%, у груши - 43%; корни длиной от 6 до 10 мм - соответственно 18,7 и 28%. Остальные корни имеют длину от 1 - 2 см до 0,5 м, но их гораздо меньше (у яблони 16,2%). Средняя длина корня у однолетних сеянцев плодовых пород довольно постоянная: у яблони 6 - 7 мм, у груши и черешни 8 - 9, у вишни 8, у антипки 14 мм. У плодоносящих деревьев плодовых пород средняя длина корня в 2 раза меньше (у яблони 3,5 мм), чем у однолетних саженцев.
Циклическая смена (самоизреживание) корней. Дю-Монсо (1760) наблюдал отмирание, или изреживание, корней в корневой системе растений, то есть освобождение ее от мелких корней и замену их новыми, а Найт (1806) заметил у многих деревьев отсутствие главного корня. Это явление объяснимо, если рассматривать развитие растительного организма как единство двух противоположных, но взаимосвязанных процессов - возникновения и отмирания.
Наши многолетние исследования позволили установить, что в каждом растении с первых дней и до конца жизни происходит неуклонное и последовательное отмирание концов осевых корней, а также коротких боковых корней сначала на главном корне, затем по мере роста на более длинных корнях последующих порядков ветвления (первом, втором и т. д.); отмирание целых мочек (сеток) корней и замена их новыми, то есть очищение (оголение) верхних, уже более толстых корней от отдельных более мелких корней и мочек (рис. 14).
На однолетних сеянцах за один сезон могут отмиреть десятки тысяч, а у взрослых деревьев - миллионы корней. Это свидетельствует о недолговечности всасывающих корней, которые в зависимости от расположения в корневой системе живут от нескольких дней до месяца и редко дольше. Отмирание корней названо нами корнепадом, оно несколько созвучно с явлением листопада у деревьев.
У разных пород отмирание корней может ускоряться или замедляться в разной степени, в зависимости от условий жизни. В результате отмирания и возобновления корневая система растения непрерывно в течение вегетации и всей жизни продвигается во все новые, неиспользованные слои почвы и почвогрунта. Она распространяется от ствола круговыми полосами, отмирая в одном месте и появляясь в другом, и обеспечивает нормальные условия роста и плодоношения плодовых растений (рис. 15). Суммарная масса отмерших в течение года корней выражается десятками килограммов и даже тоннами органических остатков в расчете на гектар, являющихся источником питания микроорганизмов ризосферы.
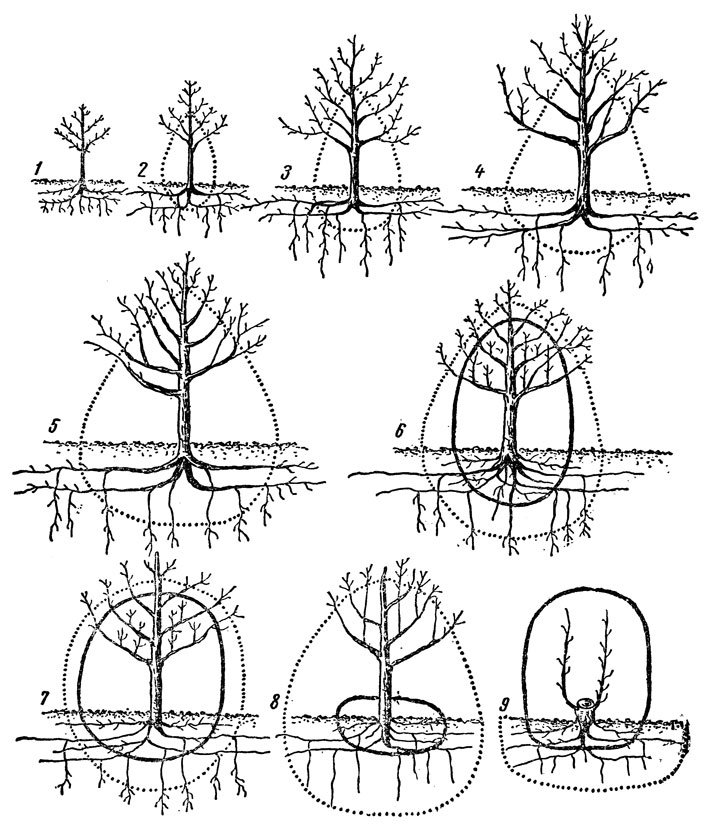
Явление отмирания корней установлено многими советскими и зарубежными учеными. Так, выявлено, что отмирают не только обрастающие (мелкие) корни, но и скелетные.
А. Д. Крестников (1961) нашел, что яблоня в старших периодах обладает способностью образовывать корни из корневой шейки сеянцевого подвоя. Следовательно, масса всасывающих корней часто находится не только за пределами проекции кроны, но и иногда около ствола, что надо учитывать при уходе за плодовыми культурами.
Отмирание и возобновление корней, или циклическая смена их, у сеянцев и взрослых плодовых и ягодных растений представляет естественный процесс в жизненном цикле их развития и, по нашему мнению, свойственно всем древесным и травянистым растениям.
Формирование корневой системы. Рост и формирование корневой системы в почве зависят от происхождения корней (семенные или вегетативные), породы и сорта растения (привой), природных условий и агротехники.
Корневые системы семенного происхождения обычно проникают в почву глубже, чем вегетативного происхождения.
Различны типы роста корней в стороны и в глубь почвы, а также их расположение по горизонтам почвы и почвогрунта. Например, в Московской области ежегодный прирост корней плодовых культур в стороны от ствола по радиусу составлял в среднем 20 - 36 см, а в глубину - 17 - 33 см; в Крыму - соответственно 20 - 25 и 5 - 15 см; на Кубани - 38 и 50 - 100 см. Различия значительны, плодовод должен учитывать их при выборе и подготовке почвы под сад и при дальнейшем уходе за корнями и почвой. Агротехническими приемами, например плантажной вспашкой, обработкой и удобрением, на любой почве, в том числе и на дерново-подзолистой, можно увеличить массу корней, расположенных ниже пахотного горизонта.
Тонкие, обрастающие и всасывающие корни в зависимости от природных условий и агротехники могут расти с ранней весны и до поздней осени, даже после листопада, а затем прекращать рост и отмирать.
В общем постепенно разрастаясь, корневая система сеянца, а позднее взрослого плодового дерева охватывает огромные объемы почвы и почвогрунта длинными и толстыми тяжами корней, на которых размещены сотни тысяч коротких корешков, а на них десятки миллионов корневых волосков.
Корни горизонтального направления у плодовых пород располагаются в северной зоне в массе на глубине примерно до 30 - 50 см, в средней до 50 - 75 см, в южной (Кубань) до 100 - 120 см. По данным Г. И. Груздева (1937), в различных зонах европейской части СССР основная масса корней плодовых пород расположена в перегнойно-аккумулятивном горизонте и в верхней части иллювиального горизонта почвы. Например, в аккумулятивном горизонте А в лесостепной почве их было 78%, в черноземной - 66, в темно-каштановой - 48, в сероземе - 33 и в дерново-подзолистой - 36%.
В плодовых садах Среднего Поволжья на тяжелосуглинистой почве основная масса корневой системы яблони была равномерно расположена в слое 0 - 90 см, а на карбонатном суглинистом черноземе больше всего корней в гумусовом горизонте (до глубины 40 см). Корневая система яблони на аллювиальных почвах Грузии размещается на глубине до 40 см (36%), на сероземах Узбекистана - 20 - 60 см, а у абрикоса - на глубине от 5 до 60 см. Корни фундука на Кубани залегали на глубине 50 - 60 см, а в Азербайджане - только до 20 см. В Крыму у ореха грецкого корни горизонтального направления находятся на глубине до 80 см, а у хурмы восточной в Грузии - от 10 до 100 см, в Азербайджане - до 10 - 20 см. На луговой суглинистой почве поймы Днестра корни яблони были расположены до. глубины 60 см (66,9%), а у сливы 60% корней находилось на глубине до 50 см.
В Московской области на дерново-подзолистых почвах основная масса корней горизонтального направления расположена у яблони на глубине до 75 см, у груши до 50, у вишни до 40, у сливы до 30 см (рис. 16).
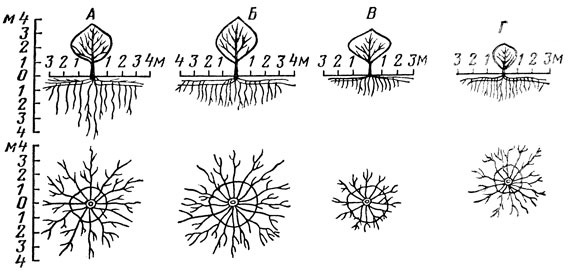
В результате наличия плотных, а местами сцементированных прослоек или погребенных горизонтов в почве корневая система залегает ярусами. Например, на Кубани первый ярус корней яблони залегает на глубине до 75 - 100 см, второй - 220 см, в Среднем Поволжье на дерново-подзолистой почве - соответственно на глубине 18 и 101 - 140 см.
Более тонкие корни в значительных количествах могут встречаться во всех зонах плодоводства в поверхностном слое почвы с глубины 3 - 10 см. У яблони всюду на поливных землях и на севере корни чаще размещены глубже 10 - 20 см от поверхности, а в засушливых районах глубже 30 - 40 см, например в совхозе "Агроном" Липецкой области.
Исследования, проведенные нами в Московской области, показали, что у 7-летних яблонь корневая система разрастается в стороны по диаметру на 3,5 м, у 14-летних - на 5 м, у 20-летних - на 8 - 9 м. Следовательно, при 8-метровом междурядье свободное от корней пространство на 7-летних посадках будет 4,5 м, на 14-летних - 3 м, на 20-летних посадках корни заполняют все междурядье и входят в другое междурядье на 0,5 м (рис. 17). В то же время даже к 20 - 22 годам разные сорта яблони сохраняют свободное пространство до 1 - 3 м между кронами деревьев соседних рядов.
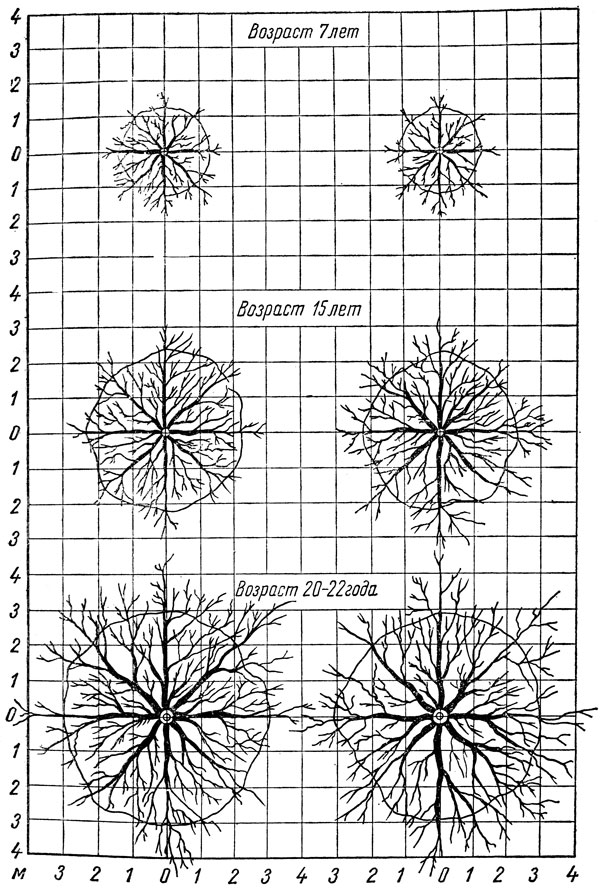
Советские и зарубежные исследователи установили, что у плодовых пород, начиная со второго года, и всю последующую жизнь диаметр корневой системы в 1,5 - 2 раза больше диаметра (проекции) кроны. Такое соотношение диаметра крон и корневых систем наблюдается у большинства пород во всех зонах возделывания.
Корни вертикального направления у разных плодовых культур проникают в северной зоне до 1 - 2 м, в средней до 2 - 4 м, а в южной (Крым, Кубань) до 5 - 9,5 м и даже до 12 м (Казахстан). Вглубь корни продвигаются главным образом по ходам. землероев, особенно дождевых червей, и по трещинам почвы.
На глубину проникновения корней сильное влияние оказывают почвенные условия, порода, привой и подвой. На дерново-подзолистой почве (Московская область) корни яблони проникают на глубину до 3 - 4 м, груши, сливы и вишни - до 2 м. На этой же почве основная масса вертикальных корней Антоновки обыкновенной на китайке залегает на глубине 1 м, на дусене III - 1,3 м, на яблоне лесной и сеянцах Аниса - 2 м, а на сеянцах Антоновки - на глубине 2,5 м.
На различных почвах Молдавской ССР корни яблони, сливы, абрикоса и айвы проникают на глубину до 2 м, корни груши - до 2,5 м, на Украине (Мелитополь) корни черешни на антипке (привой черешня) проникали на глубину более 4 м, на черешне - 3,2 м на вишне - на глубину не более 2 м.
В Крыму корни яблони на парадизке и груши на айве проникали на глубину 4 - 4,5 м, в Молдавии корни трехлетнего ореха грецкого - на глубину 5 м.
Количество и длина корней вертикального направления могут быть значительными. Например, у яблони Сары синап в возрасте 45 лет (Крым, совхоз им. Чкалова) общая длина всех скелетных и обрастающих корней равнялась 2,7 км, из них корней вертикального направления было 1,6 км, а горизонтального 1,1 км; под кроной находилось 36% корней. В Казахстане под кроной яблони было 30 - 40%, а вне ее 64% и тех и других корней; в Грузии - соответственно 38,9 и 61,1%.
У яблони Ренет шампанский в возрасте 25 лет в том же совхозе им. Чкалова, но на другом участке с близким стоянием грунтовых вод глубина проникновения корней в почву составляла только 1,3 м, а общая длина всех корней достигала 543 м; длина вертикальных корней была только 77 м, а горизонтальных - 456,6 м. В пределах кроны находилось 216,6 м корней, за ее пределами - 240 м. Количество вертикально идущих корней откопанных деревьев яблони в Крыму колебалось от 13,6 до 64,7%, а длина - от 11,2 до 58,12% всех корней дерева.
Корневые системы деревьев вишни (Московская область), выращенных из черенков, уступали корневым системам привитых деревьев по мощности отдельных скелетных корней, но обладали большей массой разветвленных мочек.
Корни плодовых деревьев при соответствующих условиях могут довольно быстро проникать вглубь. Например, при одних условиях корни сильнорослых подвоев плодовых деревьев уже в возрасте 4 - 12 лет проникают на глубину 3 - 3,5 м и даже 9,5 м (Кубань), а при других условиях корни 30-летних деревьев - только на 1 - 1,5 м (Крым). Оказалось, что корни и карликовых подвоев (райка и айва) в Крыму также могут проникать до 4 - 5,5 м в глубину.
В Крыму деревья яблони сорта Сары синап в возрасте 35 лет при размещении корней на глубине до 4,6 м давали до 300 кг плодов, а деревья того же сорта и возраста при других почвенных условиях (близкие грунтовые воды) и глубине залегания корней до 1,5 м давали только 100 кг плодов.
Как правило, чем южнее произрастают плодовые и ягодные культуры, тем глубже залегают их корневые системы. Однако и на юге корневые системы могут быть расположены поверхностно. Это наблюдается в случаях близкого залегания грунтовых вод или плотных, а иногда галечных прослоек и сцементированных отложений, особенно на аллювиальных почвах южной зоны СССР.
Лучшей корневой системой плодовых пород следует считать такую, которая залегает равномерно по кругу, более глубоко и широко и имеет возможность извлекать воду и питательные вещества из большего объема почвы и почвогрунта. Плодовое дерево в этом случае характеризуется большой засухо- и морозоустойчивостью.
Исследованиями советских и зарубежных ученых установлено, что плодовые деревья с мощной корневой системой гораздо дольше живут, более зимостойки, лучше и регулярнее плодоносят. Поэтому очень важно соответствующими агротехническими приемами способствовать глубокому залеганию корневой системы. Вместе с тем плодовод всегда перед вспашкой должен проверять глубину залегания корней, чтобы правильно назначить в каждом саду и на каждом его участке глубину вспашки, внесение удобрений и т. п.
Взаимовлияние корней. Еще Ч. Дарвин отмечал острую конкуренцию корней между разными экземплярами одного и того же вида при совместном произрастании и отсутствии ее у разноименных культур. П. Г. Шитт (1913) и Т. К. Кварацхелия (1927) установили, что корни плодовых деревьев распространяются в направлении слабого соседнего дерева.
В наших многолетних исследованиях в Крыму и на Кубани (1929, 1937 гг.) наблюдались следующие случаи взаиморасположения корней: корни яблони в одних местах не входят, а в других входят в корни соседней яблони; корни яблони входят в корневую систему выкорчеванного в прошлом году дерева яблони, но уходят от корневых систем сеянцев яблони только что выкорчеванного питомника; корни сливы, черешни свободно входят в корневую систему яблони; то же самое наблюдалось в отношении корней яблони, груши, абрикоса, миндаля и айвы, то есть между корнями этих пород нет конкуренции.
Особенно рельефно себя проявили корневые системы персика, что нами установлено на Кубани (1928) на двух соседних сеянцах 4-летнего возраста, у которых корни резко разошлись, оставив коридор шириной 80 - 120 см (рис. 18).
Такое же явление наблюдалось в Италии (Баргиони, 1959). Здесь не смыкались корни и плодоносящих деревьев персика. Позднее, сравнивая расположение корневых систем хурмы, винограда и вишни, Баргиони подметил, что корни этих пород переплетаются между собой, тогда как пересечения корней растущих близко одно к другому деревьев хурмы не наблюдалось.
По данным Н. А. Тхагушева и Л. С. Наумова (1969), у кустов фундука (до 15-летнего возраста) при площади питания 5 Х 5 м смыкания или переплетения корней не наблюдается.
Некоторые исследователи считают, что на расположение корней в однопородных или разнопородных посадках влияют почвенные условия и корневые выделения (токсины), полезные или вредные для корней соседнего дерева. Исследования А. Скиляси (1968) в Италии показали возможность совместной посадки маслины и цитрусовых только на почвах богатых, влажных и обладающих хорошими физико-механическими свойствами. На почвах, менее благоприятных, корневая система маслины уходит далеко за пределы проекции кроны и угнетает корневую систему цитрусовых, заметно снижая их продуктивность.
Т. Л. Исаева (1968) установила, что корневые выделения абрикоса задерживают появление проростков абрикоса, но стимулируют рост проростков груши. Корневые выделения груши содержат нитраты, фосфаты, соли калия и органические кислоты, аминокислот не обнаружено. Корневые выделения абрикоса содержат только нитраты и соли калия. В целом корневые выделения древесных пород играют большую роль в биохимических процессах, процессах роста и развития растений, и их влияние необходимо учитывать при создании долголетних устойчивых насаждений.